North American Bullfrog
Lithobates catesbeianus
Common Name: |
American Bullfrog |
Scientific Name: |
Lithobates catesbeianus |
Etymology: |
|
Genus: |
Lithobates is Greek, Litho means "A stone", bates means "One that walks or haunts." |
Species: |
catesbeianus is in honor of Mark Catesby (1679-1749) |
Average Length: |
3.5 - 6 in. (9 - 15.2 cm) |
Virginia Record Length: |
|
Record length: |
8 in. (20.3 cm) |
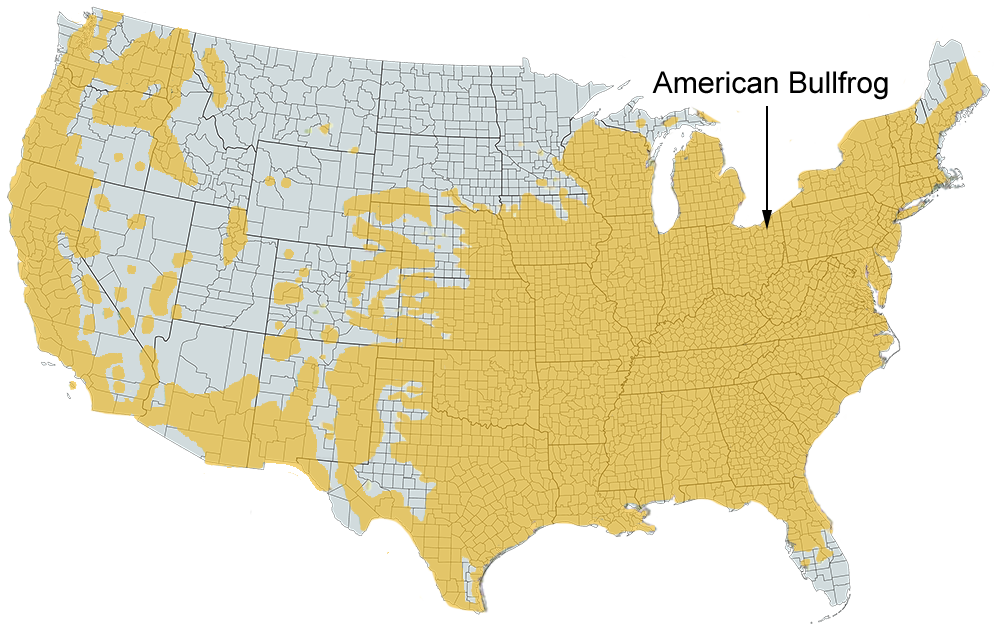
Historical versus Current Distribution - Historically, American Bullfrogs (Lithobates catesbeianus; hereafter in this account referred to as Bullfrogs) had one of the largest ranges of any North American amphibian (Bury and Whelan, 1984), occurring from Nova Scotia south to central Florida and west across the Great Plains (Conant, 1975), and probably including Tamaulipas and northern Vera Cruz in northern Mexico (Kellog, 1932). Although the historical range was large, Bullfrogs were naturally limited in distribution (Bleakney, 1958). Accurate determination of the original distribution will be forever confused as a result of introductions outside the natural range in the western United States (Bury and Whelan, 1984; see "Interspecific Association/Exclusions" below), as well as within the natural range in the Midwest (Lannoo et al., 1994; Casper, 1996; Christiansen, 1998; Hemesath, 1998; Moriarty, 1998). Bullfrogs were introduced into Oregon as early as the 1920s or 1930s (Nussbaum et al., 1983). Bullfrogs have also been introduced into Europe (Stumpel, 1992; Thiesmeier et al., 1994), South America, Asia (Kupferberg, 1997), British Columbia, Mexico, the Caribbean Islands, Brazil, and Hawaii (Bury and Whelan, 1984), due to their culinary appeal. Local extirpations have been reported in southwestern Ontario (Hecnar, 1997).
Historical versus Current Abundance - In general, frog populations fluctuate widely in response to seasonal precipitation, disease, predation, and other factors, making the discernment of overall trends problematic. Shirose and Brooks (1997) estimated that Bullfrog populations in Algonquin Park, Ontario, likely fluctuate by as much as 50–80%, even in the absence of long-term trends in population size. Nevertheless, Bullfrogs remain common throughout most of their natural range, despite general depression of frog numbers in many regions.
Historically, Bullfrogs were common in the United States and abundant despite harvesting (Baker, 1942). Today many native Bullfrog populations appear to be declining, with habitat loss and degradation, water pollution, and pesticide contamination (see "Conservation" below) commonly invoked as causal factors (Texas, Baker, 1942; Louisiana, Walker, 1963; United States, Bury and Whelan, 1984; Great Lakes, Harding, 1997; Wisconsin, Mossman et al., 1998). Wetland drainage, shoreline development, and damage to the wetland fringes of lakes from home building and recreation have greatly decreased Bullfrog habitat quality and availability in many areas (Bury and Whelan, 1984; Hunter et al., 1992; Casper, 1998). Over-harvesting has also been implicated in Bullfrog decline (Baker, 1942; Gibbs et al., 1971; Treanor and Nichola, 1972; Treanor, 1975; Vogt, 1981).
The creation of farm and golf course ponds, where high Bullfrog densities can develop in altered wetland habitats, has increased Bullfrog abundance in some areas (Willis et al., 1956; personal observation, 1990s), and widespread introductions have greatly expanded the range. Introduced populations appear to be experiencing range expansions in Iowa (+ 279% increase; Christiansen, 1998), Minnesota (Moriarty, 1998), and California (Bury and Luckenbach, 1976), although Treanor and Nichola (1972) report declines in the late 1960s in California.
Breeding - Reproduction is aquatic.
Breeding migrations - Breeding takes place in spring and early summer and is earliest in southern latitudes (Bury and Whelan, 1984). In Texas, Bullfrogs breed from March–October (Blair, 1961a); in Québec, breeding occurs from late May to mid July (Bruneau and Magnin, 1980); and in the Great Lakes in June–July (Pentecost and Vogt, 1976; Harding, 1997; Mossman et al., 1998; Varhegyi et al., 1998). Lepage et al. (1997) provides phenology data for Québec, with the peak calling period occurring from mid June to July. However, Bishop et al. (1997) give 17 May to 8 June as peak calling dates in southern Ontario. In Kansas, calling begins when air temperatures are above 21 ˚C (Fitch, 1956c). In Michigan (and probably elsewhere, especially in southern latitudes), double clutching occurs, with some females active early and then again 3 wk later (Emlen, 1977). Older females may produce two clutches/year, with second clutches containing substantially smaller eggs (Howard, 1978b).
Breeding habitat - Bullfrogs breed in the vegetation choked shallows (Pope, 1964a) of permanent bodies of water. Male Bullfrogs will aggressively defend oviposition sites (Ryan, 1980) by pushing, shoving, and biting. Females select a mate by entering his territory (Ryan, 1980). Female mate choice becomes more discriminating with age, with older females consistently selecting the oldest, largest males as mates (Howard, 1978a). Older females sometimes vocalize within male choruses, which may elicit higher levels of male–male competition and assist females in selecting high-quality males (Judge et al., 2000). Embryo mortality depends on female choice of the oviposition site, the best of which are controlled by the largest males and where water temperatures do not exceed 32 ˚C, over which developmental abnormalities occur (Howard, 1978b). Howard (1979) estimated male reproductive success in a Michigan population. The most successful males fertilized three females, the least successful had no matings, and the average was 0.71 matings/male. Males defending territories in the middle of ponds are susceptible to predation by the snapping turtle, Chelydra serpentina (Emlen, 1976; Howard, 1978a).
Egg deposition sites - Eggs are laid in thin sheets on the water surface, covering 0.5–1 m2, and hatching in 3–5 d (Bury and Whelan, 1984).
Clutch size - Bullfrogs are extremely prolific, producing up to 20,000 eggs/clutch (Schwalbe and Rosen, 1999). The number of zygotes sired by successfully mating males ranged from 4,928–59,035 (mean 11,149), with the number of resultant hatchlings ranging from 299–29,377 (mean 5,582; Howard, 1978b). Females may lose up to 27% of their body mass during oviposition (Judge et al., 2000).
Altig & McDiarmid 2015 - Classification and Description:
- Eastern Film
- Arrangement 3 - Film diameter greater than 200 mm; ovum black and jelly diameter not
large relative to ovum size.
- Sub-arrangement A - Eggs deposited in larger nonflowing water; film greater than 150 mm diameter; 1 jelly layer; Ovum Diameter 1.2-1.7 mm; Egg Diameter 6.4-10.4 mm; clutch 10,000-20,000;
- Arrangement 3 - Film diameter greater than 200 mm; ovum black and jelly diameter not
large relative to ovum size.
Larvae/Metamorphosis - Tadpoles favor warm water environments (24–30 ˚C; Brattstrom, 1962b) and have preferred body temperatures ranging from 15–26.7 ˚C, which changes with acclimation (exposure to new temperature ranges), developmental stage, and season (Ultsch et al., 1999). Oxygen consumption increases about exponentially as a function of temperature (Burggren et al., 1983; Feder, 1985).
Tadpoles:

| Lateral View | Dorsal View |
|---|---|
| BL = Body Length | IND = Internarial Distance |
| MTH = Maximum Tail Height | IOD = Interorbital Distance |
| TAL = Tail Length | TMW = Tail Muscle Width |
| TL = Total Length | |
| TMH = Tail Muscle Height |

Length of larval stage - The time to metamorphosis varies from a few months in the south to 3 yr in Michigan and Nova Scotia (Collins, 1979; Bury and Whelan, 1984). The length of the larval period is negatively correlated with mean length of the frost-free period, and tadpoles in a Michigan study transformed in either the first or second season following the hatching year (Collins, 1979; see also Crawshaw et al., 1992).
Larvae Food - The diet of larvae consists mainly of algae, aquatic plant material, and some invertebrates (Treanor and Nichola, 1972; Bury and Whelan, 1984). Growth rates and digestive abilities of tadpoles fed differing algal diets was studied by Pryor (2003). Conflicting reports exist as to whether larvae will scavenge dead animal material (Dickerson, 1906; Funkhouser, 1976). Coprophagy was demonstrated by Steinwascher (1978a), who suggested that this increased the amount of time food was resident in the digestive tract and may also allow some microbial digestion.
Cover - Tadpoles are solitary (Punzo, 1992b) and move into deeper water until just prior to metamorphosis, at which time their depth preference is reversed (Goodyear and Altig, 1971; Smith, 1999). Bullfrog tadpoles are bimodal breathers (Crowder et al., 1998; Ultsch et al., 1999) and their distribution within ponds is likely influenced by both temperature and oxygen availability in addition to predator avoidance (Nie et al., 1999). Bullfrog tadpoles avoid predators by seeking cover (e.g., Pearl et al., 2003).
Larval polymorphisms - Not reported.
Features of metamorphosis - Metampophosis is asynchronous. Newly metamorphosed animals generally frequent shorelines where vegetative cover is thick enough to afford protection from predators (G.S.C., personal observation). Thrall (1971) found newly metamorphosed Bullfrogs occupying small circular pits along bare mud flats of a farm pond in Illinois.
Post-metamorphic migrations - Specific information is lacking
Juvenile Habitat - Bullfrogs are adept at colonizing newly created ponds (Merovich and Howard, 2000) and the juvenile stage is likely where most dispersal occurs.
Adult Habitat - Adult Bullfrogs are mostly aquatic and prefer warmer, lentic habitats (George, 1940) such as vegetated shoals, sluggish backwaters and oxbows, farm ponds, reservoirs, marshes, and still waters with dead woody debris (Holbrook, 1842; Storer, 1922) and dense and often emergent vegetation (Bury and Whelan, 1984). They also occupy shorelines of lakes and streams. In a long-term study of ponds in southern Michigan, Skelly et al. (1999) found Bullfrogs restricted to open canopy ponds with permanent water. Bullfrogs prefer relatively high body temperatures (26–33 ˚C; Lillywhite, 1970) and actively thermoregulate, basking and posturing to control body temperatures (Lillywhite, 1970, 1971, 1974b). Adults generally remain inactive until water temperatures approach 15 ˚C (Harding, 1997). Unlike many other frogs, Bullfrogs can coexist with predatory fishes (Hecnar, 1997).
Home Range Size - Currie and Bellis (1969) found Bullfrogs to have a mean activity radius of 2.6 meters in an Ontario pond. Their data suggest that home range size is reduced at higher densities and increases as the size of an individual increases, and that males have a larger home range than females. Raney (1940) found no evidence of homing in a New York population. Dispersals of at least 3.2 km from the home pond have been recorded, with dispersal distances of 7–8 km likely (Schwalbe and Rosen, 1999). Maximum movement distances of up to 1,600 m (mean 402), as well as homing, were reported by Ingram and Raney (1943) in New York. McAtee (1921) noted a Bullfrog that returned to its lakeshore territory within 2 d after being moved ≤ 0.4 km away. Schwalbe and Rosen (1999) reported an adult Bullfrog every 1.8 m along some shorelines in Arizona ponds. In Kentucky, Cecil and Just (1979) calculated densities of 0.9–13.2 tadpoles/m2 of pond, biomass of tadpoles as 11–103 g/m2 of pond, and survival rates of 11.8%, 13.1%, and 17.6%. Treanor (1975) reports densities in California canals from 6.6–119 frogs/km, with average numbers reduced by 20–25% in areas where harvest occurred. Bullfrog density in an Ontario pond was 0.9 and 1.3/m2 (Currie and Bellis, 1969) and 8.8–45.8 frogs/ha in a 7 ha Illinois lake (Durham and Bennett, 1963).
Territories - Bullfrogs are highly territorial and have a polygynous mating system, with the largest males in a population controlling the highest quality oviposition sites preferred by females (Howard, 1978a,b). Males defend roughly circular territories 2–5 meters in diameter (Harding, 1997). Bullfrogs recognize the calls of neighbors and respond more strongly to the calls of strangers (Davis, 1987). Most physical encounters are won by larger males by engaging in wrestling, shoving, and pouncing (A.P. Blair, 1963; Durham and Bennet, 1963). Younger males unable to hold territories against larger, older males, employ male parasitism (intercepting females attracted to large males) and opportunism (temporarily holding a vacated territory until threatened by a larger male) as mating strategies (Howard, 1978a). In an Illinois population, Mauger (1988) observed medium-sized males engage in opportunism, where they vocalized from the fringes of a large male territory until forced to flee or they inhabited recently vacated territories. The smallest males practiced parasitism, maintaining a low posture without vocalizing in an attempt to intercept females en route to a large-male territory. Males near a large-male territory also invade such territory when the large male is amplexed, without challenging or displacing the resident male.
Aestivation/Avoiding Dessication - No information regarding aestivation has been reported.
Seasonal Migrations - Bullfrogs are the last North American ranid to emerge in spring (Voice, 1931; Smith, 1934; Ryan, 1953), following a general northward progression (Bury and Whelan, 1984). However, Bullfrogs in Louisiana may be active throughout the year (George, 1940). Overland movements, such as the dispersal of newly metamorphosed animals and adult movements, occur on warm, rainy nights (Raney, 1940; Gibbons and Bennett, 1974; Johnson, 1987; G.S.C., personal observation). Summer movements appear to be limited to adverse or drought conditions (Jameson, 1956a). In Missouri, only a small percentage of frogs migrated between ponds, and the few that did traveled 0.16–2.8 km (Willis et al., 1956).
Torpor (Hibernation) - Bullfrogs are not freeze tolerant. In autumn, adults become torpid before frost occurs, while juvenile activity continues until freezing weather (Willis et al., 1956). Winter hibernation usually takes place under water (Treanor and Nichola, 1972), where individuals bury themselves in surface mud or construct protective pits (Emery et al., 1972; Johnson, 1987; Harding, 1997) or cave-like holes (Ruffner, 1933). Bohnsack (1952) reported a torpid Bullfrog from a terrestrial hibernaculum, buried under leaf litter in a soil pocket of an oak-hickory woods in Michigan. Radio-tagged Bullfrogs in Nova Scotia hibernated in a pond and tolerated prolonged hypoxia (sustained PO2 below 20 Torr for > 8 wk; Friet and Pinder, 1990). In a radio telemetry study in Ohio, Bullfrogs overwintered in relatively shallow areas (< 1 m) near small inlet streams of ponds, as well as 1–2 m off-shore of ponds, and remained active throughout the hibernation period (Stinner et al., 1994). These Bullfrogs laid on the pond bottom and were not covered by silt. Juvenile Bullfrogs cease motor activity between 0–1 ˚C (Lotshaw, 1977).
Interspecific Associations/Exclusions - Bullfrogs are often the predominant frog species in permanent aquatic habitats, naturally occurring with species such as Green Frogs (Lithobates clamitans; Werner, 1991). Introduced Bullfrogs have been implicated in the decline or displacement of many amphibians and a few reptiles, including Mexican Gartersnakes (Thamnophis eques), lowland leopard frogs (Lithobates yavapaiensis), and Chiricahua Leopard Frogs (Lithobates chiricahuensis; Arizona, Schwalbe and Rosen, 1988); Foothill Yellow-legged Frogs (Rana boylii; California, Kupferberg, 1997); Northern Red-legged Frogs (Rana aurora; Oregon, Kiesecker and Blaustein, 1997a, 1998); the apparently now extinct Vegas Valley Leopard Frog (Lithobates fisheri; Nevada, Moyle, 1973; Cohen, 1975); Northern Leopard Frogs and Plains Leopard Frogs (Lithobates blairi; Colorado, Hammerson, 1982a,b); Northern Leopard Frogs and Oregon Spotted Frogs (Rana pretiosa; Montana, Black, 1969a); Oregon Spotted Frogs (Dumas, 1966); Blanchard’s Cricket Frogs (Acris crepitans blanchardi; Ontario, Oldham, 1992); Pacific Treefrogs (Dryophytes regilla; Oregon, Jameson, 1956b; California, Kupferberg, 1997); and native amphibians in Iowa (Lannoo et al., 1994). Schwalbe and Rosen (1988, 1999) believe Bullfrogs to be responsible for declines in Mexican Gartersnakes from the San Bernardino National Wildlife Refuge and present supporting evidence.
An analysis of multiple factors correlated with the decline of northern red-legged frogs in western Washington did not support the hypothesis that Bullfrogs were excluding this species, implicating instead the presence of exotic fishes (Adams, 1999). In controlled enclosure experiments testing survival of northern red-legged frog and Pacific treefrog larvae, the presence of sunfish (Centrarchidae) reduced survivorship of both species to near zero, while the presence of Bullfrog tadpoles had a weak survival effect only in northern red-legged frogs (Adams, 2000). In another enclosure experiment, testing the effects of fishes and Bullfrogs on northern red-legged frogs, Kiesecker and Blaustein (1998) found that time to metamorphosis increased and mass at metamorphosis decreased when northern red-legged frog tadpoles were exposed to either larval or adult Bullfrogs.
In Kupferberg’s (1997) enclosure experiments, the presence of Bullfrog tadpoles resulted in a 48% reduction of survivorship of foothill yellow-legged frog tadpoles, and a 24% decline in mass at metamorphosis. Kupferberg (1997) also found a 16% reduction in the size of newly metamorphosed Pacific treefrogs, but no significant effect on survivorship.
Fisher and Shaffer (1996) found a negative correlation between the presence of introduced exotics (Bullfrogs and fishes) and native amphibians in California. However, they did not discriminate between fishes and Bullfrogs in their analyses. Hayes and Jennings (1986) argued that while Bullfrogs are often invoked as being responsible for declines in native ranid frogs in western North America, existing data cannot distinguish adequately among three major causal hypotheses for this decline: Bullfrogs, habitat alteration, and introduced predatory fish. They believed that fish predation was the most compelling hypothesis.
In Ontario, Hecnar and M’Closkey (1997) observed the relative abundance of green frogs increase fourfold after Bullfrog extirpation, where competition between these species and predation of Bullfrogs upon green frogs is postulated. In a study of Bullfrog/green frog interactions in Michigan, Werner et al. (1995) concluded that Bullfrogs relied more heavily on aquatic prey than did green frogs, that overlap between the species in diet resources declined with increasing body size disparity, and that juvenile frogs were a major component of the diet of Bullfrogs but not green frogs. Stewart and Sandison (1972) found both dietary and habitat differences, with some overlap, in sympatric populations of Bullfrogs, mink frogs (Rana septentrionalis), and green frogs in New York. Werner (1991) demonstrated that small larvae of Bullfrogs and green frogs reduced activity in the presence of predatory Anax dragonfly naiads, but Bullfrogs did less so, thereby gaining a competitive advantage over green frogs when naiads were present. This was a size-dependent effect, as larger Bullfrog tadpoles largely ignored the presence of naiads.
In pair-wise competition experiments with eight species of desert anurans, Woodward (1982) found that Bullfrog tadpoles were the only permanent pond species able to persist in the presence of Couch’s spadefoot (Scaphiopus couchii) tadpoles, and that temporary pond species were competitively superior to species breeding in permanent waters.
In laboratory experiments, Alford (1989c) studied competition between Cope’s Gray Treefrog (Dryophytes chrysoscelis) and Bullfrog tadpoles and found that the presence of Bullfrogs substantially reduced the mass of Cope’s gray treefrogs, but that reproductive phenology influenced this effect. Peacor and Werner (1997) conducted experiments in cattle watering tanks with Bullfrog tadpoles, green frog tadpoles, adult Bullfrogs, the lethal presence of the larval odonate predator Tramea lacerata, and the nonlethal (caged) presence of the larval odonate predators Anax junius and Anax longipes. They demonstrated that the presence of large Bullfrog competitors increased the rate of Tramea predation on small green frogs and small Bullfrogs, and that the presence of nonlethal Anax increased the competitive advantage of Bullfrogs over green frogs. The proposed mechanisms were changes in small tadpole activity.
Moore (1952) attributed the southern distributional limit of mink frogs to competitive exclusion by Bullfrogs. In sum, interactions among Bullfrogs, other amphibians, and the environment is complex and is influenced by both the predators and resources present in the system, as well as the phenology of life history events.
Age/Size at Reproductive Maturity - Sexual maturity in Bullfrogs usually occurs in 1–2 yr in males, and 2–3 yr in females (Howard, 1981), with males maturing at an earlier age and/or smaller size than females. Turner (1960) reported maturity in 1 yr; Raney and Ingram (1941) reported 2–3 yr in a New York population; Minton (1972) gives 1 yr after metamorphosis; and Carr et al. (1984) recorded ovarian maturity in females 12 mo after metamorphosis under lab conditions. Harding (1997) reports Bullfrogs reaching breeding size in 2–4 yr when they are 10–12 cm.
The size reached at sexual maturity is influenced by geographic location (Bury and Whelan, 1984) and occurs at 95–110 mm in Québec (Bruneau and Magnin, 1980) and 125 mm or greater for females in Missouri (Willis et al., 1956). Growth rates of lab-raised juveniles slow from 27 to 14 to 8 mm/yr in the first, second, and third years, respectively (Jorgensen, 1992). The growth data of Schroeder and Baskett (1968) are summarized in Table 9. Minimum snout-ischium length of reproductive individuals in Michigan was 95 mm for males, and 108 mm for females (Howard, 1981).
Longevity - Longevity for wild Bullfrogs is estimated to be 8–10 yr, although a captive specimen survived for nearly 16 yr (Oliver, 1955a; Goin and Goin, 1962). A maximum age of > 6 yr was revealed by osteology in Bullfrogs from Missouri (Schroeder and Baskett, 1968). Survival rates of larvae are largely unknown but were reported as 11.8%, 13.1%, and 17.6% in Kentucky, where predation was the controlling factor (Cecil and Just, 1979). Larval survival rates are subject to wide yearly fluctuations in north temperate climates (Turner, 1962).
Feeding Behavior - Adult Bullfrogs are voracious, opportunistic predators (Schwalbe and Rosen, 1988) that employ a sit-and-wait approach and will readily attack any live animal smaller than themselves, including conspecifics and other frogs (Bury and Whelan, 1984). Bullfrogs may locate and eat smaller frogs by orienting to breeding (Green and Pauley, 1987; Harding, 1997) or distress (Collins and Collins, 1991) calls. Cannibalism is well documented (Bury and Whelan, 1984), and conspecifics can comprise up to 80% of the diet (Stuart and Painter, 1993). An instance of scavenging on a road-killed plains leopard frog was reported by Guarisco (1985). Frost (1935) performed stomach dissections and found that smaller Bullfrogs eat mostly insects, while larger Bullfrogs typically eat frogs, crayfish, and mice. In Arizona, the majority of stomach contents of adults consisted of invertebrates such as snails and insects (Schwalbe and Rosen, 1988), while two other studies showed 80% and 87% of stomach contents by volume to be conspecifics, mainly larvae and newly metamorphosed animals (Kansas, Smith, 1977; New Mexico, Stuart and Painter, 1993). In a New York Bullfrog population sympatric with mink frogs and green frogs, the diet of Bullfrogs differed from the other species by having fewer food types and by small ranids comprising the largest part of the diet (Stewart and Sandison, 1972). While Bullfrogs are known to consume toads (Bufo sp.), the extent of their immunity to the toxic parotoid gland secretions is debatable (Tucker, 1994). Items included in the diet of adult Bullfrogs are summarized in Table 10.
Predators - Bullfrogs are an important food source for other wildlife, often providing an abundant and localized source of protein. In many states and provinces, Bullfrogs are considered a game species and hunted for their legs. In Kentucky, Cecil and Just (1979) believed predation was the major factor controlling Bullfrog population size in permanent ponds. Bullfrog tadpoles are relatively immune to fish predation because of unpalatability (Walters, 1975; Kruse and Francis, 1977; Nelson, 1980; Kats et al., 1988; Werner and McPeek, 1994) and are one of only a few species likely to persist after fish invasion (Seale, 1980). Typically, higher rates of activity make Bullfrog tadpoles more vulnerable to invertebrate and salamander predation in fishless habitats (Werner and McPeek, 1994; Skelly et al., 1999). Table 11 lists known predators of Bullfrogs. Large males suffer higher mortality than large females (Howard, 1981). A list of prey items taken by introduced Bullfrogs in southern California has recently been published by Hovey and Bergen (2003).
Anti-Predator Mechanisms - Eggs and larvae avoid predation by fish and some salamanders by being unpalatable (see "Predators" above) or reduced larval activity (Woodward, 1983). Hobson et al. (1967) reported that adults have a state of rest characterized by alertness without a loss of reactivity, aiding in predator avoidance. Upon disturbance, adults retreat to deeper water with a series of long leaps with a great deal of splashing (Smith, 1961). Bullfrogs usually squawk when fleeing, often setting off a mass Bullfrog exodus from a shoreline into deeper waters (Schwalbe and Rosen, 1999). Bullfrogs may emit a piercing scream when seized, which may startle a predator enough to allow escape (Harding, 1997). Partial resistance to the venoms of cottonmouths (Agkistrodon piscivorus piscivorus) and copperheads (Agkistrodon contortrix contortrix) has also been reported (Heatwole et al., 1999). Bullfrog tadpoles avoid novel predators by seeking shelter, but ignore predators from their native range (Pearl et al., 2003).
Diseases - Bullfrogs are known hosts to many bacteria and viruses, but only a few of these pathogens appear to be of major importance in nature (Reichenbach-Klinke and Elkan, 1965; Carr et al., 1976; Gruia-Gray et al., 1989; Gruia-Gray and Desser, 1992; Faeh et al., 1998). An intraerythrocytic virus outbreak was reported in Canada (Crawshaw, 1997). Fungi are reported as well (Goodchild, 1953; Hill and Parnell, 1996), including a recent outbreak of chytrid fungus in Arizona (Sredl et al., 2000), which has been implicated as the cause of amphibian declines in Australia and Panama (Berger et al., 1998). In captivity, infections of Trypanosoma pipientis (Siddal and Desser, 1992), frog septicemia (Glorioso et al., 1974), Lucké tumor herpes virus (National Academy of Sciences, 1974), and polyhedral cytoplasmic amphibian virus (Faeh et al., 1998) are reported. Lefcort and Eiger (1993) demonstrated how physiological responses to pathogen infections in tadpoles (i.e., fever, malaise) can lead to increased predation. In Kentucky, Cecil and Just (1979) considered microbial infections as a secondary factor controlling Bullfrog population size in permanent ponds. Disease can be an important factor in tadpole survival (Gibbs et al., 1971).
Parasites - Bullfrogs host many parasites including helminths (Bursey and DeWolf, 1998; McAlpine and Burt, 1998), platyhelminths such as trematodes (Kennedy, 1980; Shields, 1987; Crawshaw, 1997) and nematodes (Modzelewski and Culley, 1974), protozoans and mesozoans (Bury and Whelan, 1984; Desser et al., 1986, 1990; Desser, 1987; Chen and Desser, 1989; Desser et al., 1995; Smith et al., 1996, 2000), and leeches (Barta and Desser, 1984; Siddall and Desser, 1992). Trematode metacercariae have been implicated in Bullfrog limb abnormalities (Christiansen and Feltman, 2000).
Conservation - Historically, American Bullfrogs were common in the United States. Today many native Bullfrog populations appear to be declining, with habitat loss and degradation, water pollution, pesticide contamination, and over-harvesting commonly invoked as causal factors (Texas, Baker, 1942; Louisiana, Walker, 1963; United States, Bury and Whelan, 1984; Great Lakes, Harding, 1997; Wisconsin, Mossman et al., 1998). Recent increases in the incidence of malformations (or the causal factors of same) may also be contributing to declines (Lopez and Maxson, 1990; Ouellet et al., 1997a; Rowe et al., 1998a). Exposure to pollutants (including the trace elements Arsenic, Cadmium, Selenium, Copper, Chromium, and Vanadium) in early life stages has been shown to increase the incidence of malformations, compromise swimming performance, and elevate maintenance costs (Raimondo et al., 1998; Rowe et al., 1998b; Hopkins et al., 2000). Berrill et al. (1997b) studied the effects of a variety of pesticides and herbicides on the embryos and larvae of five anuran and one salamander species. They found that Bullfrogs were the most sensitive species to fenitrothion, but that Bullfrogs in general exhibited moderate sensitivity to most contaminants, with wood frogs (Rana sylvatica) and American toads (Bufo americanus) most tolerant, and spotted salamanders (Ambystoma maculatum) least tolerant. Lethal pH (100% mortality) for both embryos and larvae is reported as 3.5–3.8, with a critical pH (where survivorship declines below levels found for neutral water) of 4.0–4.5 for embryos and 4.0 for larvae (Grant and Licht, 1993). Nixdorf et al. (1997) studied the toxic effects of lead on the central nervous system of Bullfrog tadpoles, demonstrating learning impairment. In experiments exposing Bullfrog and northern leopard frog (Rana pipiens) tadpoles to the organochlorine insecticide DDT, tail regeneration and mortality rates were less in Bullfrog tadpoles (Weis, 1975). Bullfrogs are also surprisingly resistant to the anticholinesterase drug, Sarin (isopropyl methyl phosphonofluoridate), with an LD50 of 6 mg/kg body weight (Wilber, 1954). Diana and Beasley (1998) review effects of various toxins on Bullfrogs and other amphibians. Because Bullfrog eggs are laid in thin sheets at the water surface, they are more exposed to potentially damaging solar UV-B radiation (Ovaska, 1997), which may result in embryonic mortality and abnormal development (Zimskind and Schisgall, 1955).
Widespread introductions have greatly expanded the range. Introduced populations appear to be experiencing range expansions in Iowa (Christiansen, 1998), Minnesota (Moriarty, 1998), and California (Bury and Luckenbach, 1976), although Treanor and Nichola (1972) report declines in the late 1960s in California.
American Bullfrog conservation issues vary by region. In the eastern United States, where Bullfrogs are native, attempts should be made to conserve populations by halting the causes of their declines (see above). In the western United States, efforts should be made to reduce or eliminate introduced/invasive populations.
References for Life History
- Altig, Ronald & McDiarmid, Roy W. 2015. Handbook of Larval Amphibians of the United States and Canada. Cornell University Press, Ithaca, NY. 341 pages.
- AmphibiaWeb. 2020. University of California, Berkeley, CA, USA.
- Conant, Roger and, Collins, John T., 2016, Peterson Field Guide: Reptiles and Amphibians, Eastern and Central North America, 494 pgs., Houghton Mifflin Company., New York
- Duellman, William E. and, Trueb, Linda, 1986, Biology of Amphibians, 671 pgs., The Johns Hopkins University Press, Baltimore
- Martof, B.S., Palmer, W.M., Bailey, J.R., Harrison, III J.R., 1980, Amphibians and Reptiles of the Carolinas and Virginia, 264 pgs., UNC Press, Chapel Hill, NC
- Wilson, L.A., 1995, Land manager's guide to the amphibians and reptiles of the South, 360 pp. pgs., The Nature Conservancy, Southeastern Region, Chapel Hill, NC
Photos:
*Click on a thumbnail for a larger version.




1003.JPG)




Verified County/City Occurrence in Virginia
Albemarle County
Alexandria City
Alleghany County
Amelia County
Amherst County
Appomattox County
Arlington County
Augusta County
Bath County
Bedford County
Bland County
Botetourt County
Brunswick County
Buchanan County
Buckingham County
Campbell County
Caroline County
Carroll County
Charles City County
Charlotte County
Charlottesville City
Chesapeake City
Chesterfield County
Clarke County
Colonial Heights City
Craig County
Culpeper County
Cumberland County
Dickenson County
Dinwiddie County
Fairfax County
Fauquier County
Floyd County
Franklin County
Frederick County
Fredericksburg City
Giles County
Gloucester County
Goochland County
Greene County
Greensville County
Halifax County
Hampton City
Hanover County
Harrisonburg City
Henrico County
Henry County
Isle of Wight County
James City County
King and Queen County
King George County
King William County
Lancaster County
Lee County
Loudoun County
Louisa County
Lunenburg County
Lynchburg City
Madison County
Mathews County
Mecklenburg County
Middlesex County
Montgomery County
Nelson County
New Kent County
Newport News City
Northampton County
Nottoway County
Orange County
Page County
Patrick County
Petersburg City
Pittsylvania County
Portsmouth City
Powhatan County
Prince Edward County
Prince George County
Prince William County
Pulaski County
Rappahannock County
Richmond City
Richmond County
Roanoke City
Roanoke County
Rockbridge County
Rockingham County
Russell County
Salem City
Scott County
Shenandoah County
Smyth County
Southampton County
Stafford County
Suffolk City
Surry County
Sussex County
Tazewell County
Virginia Beach City
Warren County
Westmoreland County
Williamsburg City
Winchester City
Wise County
Wythe County
York County
Verified in 106 Counties/Cities.
U.S. Range

Box Turtle Reporting
