Marbled Salamander
Ambystoma opacum
Common Name: |
Marbled Salamander |
Scientific Name: |
Ambystoma opacum |
Etymology: |
|
Genus: |
Amby is Greek for "a cup", stoma is Greek for "a mouth" |
Species: |
opacum is Latin for "shaded" or "dull". |
Average Length: |
3.5 - 4.3 in. (9 - 10.7 cm) |
Virginia Record Length: |
|
Record length: |
5 in. (12.7 cm) |
Physical Description - This species is a deep lustrous black, with brownish tinges on the underside of the head and in the legs and toes. Above they are marked with 4-7 silvery-gray (female) or white (male) cross bands that are narrow dorsally and widened on the upper sides, where they sometimes unite to endclose a series of large, fairly regular black spots which extend along the mid-line of the trunk. Costal grooves usually number 11. The length is up to 120mm.
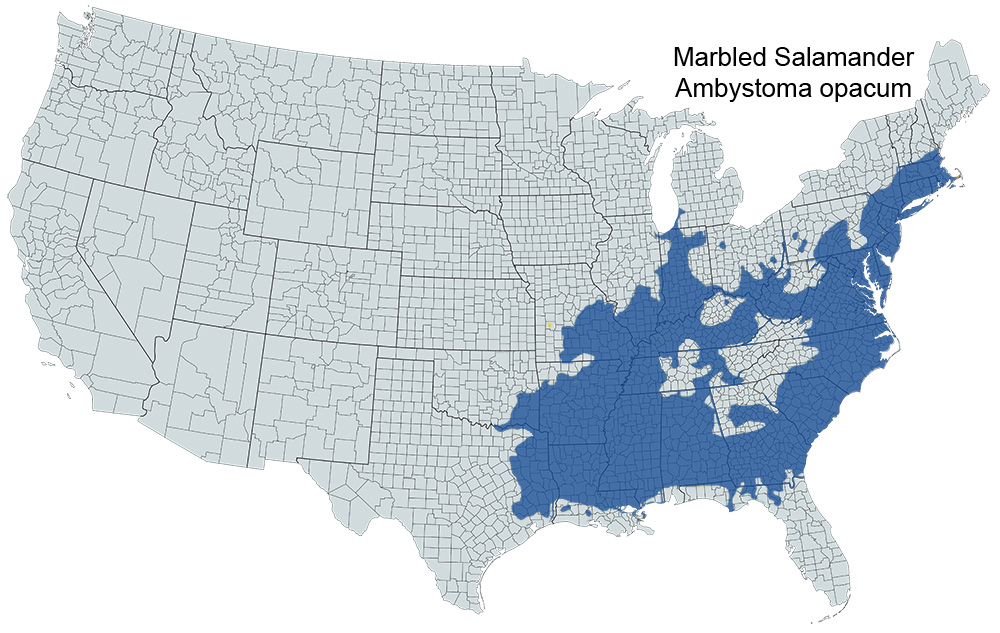
Historical versus Current Distribution - Marbled salamanders (Ambystoma opacum) range throughout much of the eastern United States from eastern Texas and Oklahoma, northeast through Illinois and Indiana to southern New Hampshire and central Massachusetts, and south to north Florida. Disjunct populations occur along the southern edge of Lake Michigan; locality data are summarized by Anderson (1967b). Additional localities are reported for east Texas (Baldauf and Truett, 1964), Louisiana (Dundee and Rossman, 1989), southeastern Oklahoma (Trowbridge, 1937), Missouri (Johnson, 1987), Mississippi (Ferguson, 1961b), Indiana (LaPointe, 1953), Alabama (Mount, 1975), north Georgia (Martof, 1955), North Carolina (E.E. Brown, 1992), and Rhode Island (Doty, 1978).
Historical versus Current Abundance - Early accounts of Marbled Salamanders describe them as common but secretive (e.g., Noble and Brady, 1933), although prior to the early 1960s no data on population sizes were collected (see Murphy, 1962; Graham, 1971). Currently, Marbled Salamanders are common and may be locally abundant in some areas. Population sizes range from dozens of individuals (Murphy, 1962) to hundreds (Graham, 1971; Shoop and Doty, 1972; Stenhouse, 1985a), ~1,000 (Pechmann et al., 1991; Semlitsch et al., 1996) to > 10,000 (Taylor and Scott, 1997). However, given the reliance of Marbled Salamanders on small isolated seasonal wetlands and intact forested floodplain habitats, their abundance presumably has declined as wetland habitats have been destroyed (Petranka, 1998). For example, from the 1950s–'70s the loss of wetlands in the Southeast was greater than in any other region of the country, with a net annual loss of 386,000 ac/yr (Hefner and Brown, 1985); in North Carolina approximately 51% of all wetland acreage on the Coastal Plain has been lost (Richardson, 1991), including 70% of the pocosins that have been “developed” or partially altered (Richardson, 1983); approximately 97% of the Carolina bays on the Coastal Plain of South Carolina have been altered or severely impacted; and < 200 bays of the original thousands are “relatively unimpacted” (Bennett and Nelson, 1991).
Breeding - Reproduction is terrestrial, in or near the wetland breeding sites prior to pond filling.
Breeding migrations - Onset of breeding migrations occurs from September–November. Timing varies geographically and may occur 1 mo or more earlier at southern latitudes compared to northern latitudes (Anderson and Williamson, 1973). On a broad scale, seasonal migrations are probably linked to regional climatic and hydrological cycles (Salthe and Mecham, 1974). Adult salamanders move to breeding sites on rainy nights and tend to enter and exit the site at approximately the same point (Shoop and Doty, 1972; P.K. Williams, 1973; Douglas and Monroe, 1981; Stenhouse, 1985a). Males generally arrive at the breeding site before females (Noble and Brady, 1933; Graham, 1971; Krenz and Scott, 1994). In a 25-yr study at Rainbow Bay in South Carolina (see Semlitsch et al., 1996), the mean date of arrival of males at the breeding site was 10 d earlier than females (unpublished data), perhaps due solely to the combination of a limited number of nights suitable for migration and slower nightly movements by fat, gravid females (Blanchard, 1930; personal observations). The sex ratio of the breeding population is often biased towards males (Graham, 1971; Stenhouse, 1987; Krenz and Scott, 1994), in part because males mature at an earlier age (Scott, 1994; Pechmann, 1994). The sex ratio in one study (Parmelee, 1993) during the non-breeding season did not differ from 1:1.
Courtship activity - At the time of autumn migration, males are at maximal testosterone levels (Houck et al., 1996; unpublished data). Courtship is terrestrial, with males exhibiting nudging, head-swinging, lifting, and body-flexing behaviors (Arnold, 1972). Spermatophore deposition follows lateral undulations of the tail. Spermatophores are 4–5.5 mm tall (Lantz, 1930; illustrated in Noble and Brady, 1933). Typical and secondary spermatophore deposition may occur (Arnold, 1972, 1976; personal observation); a male may deposit over 10 spermatophores in 30–45 min (L. Houck, personal communication). Males will mate with females outside what is typically thought of as the wetland margin (Krenz and Scott, 1994). Males often will court other males (Noble and Brady, 1933), including spermatophore deposition in the absence of a female (L. Houck, personal communication). Females may follow a male to pick up a spermatophore (Noble and Brady, 1933) or simply move about an area until a spermatophore is located (Arnold, 1972). When a spermatophore contacts a female’s vent she will lower herself onto it and insert it into her cloaca (Arnold, 1972). Sperm are stored in exocrine glands called spermathecae in the roof of the cloaca (Sever and Kloepfer, 1993). Eggs are fertilized internally by sperm released from spermathecae during oviposition (Sever et al., 1995). Females may pick up multiple spermatophores (Arnold, 1972), but sperm competition has not been definitively demonstrated. Sperm in the spermathecae do not persist for > 6 mo after oviposition (Sever et al., 1995). Males will mate with females beyond what humans typically define as the wetland margin (Krenz and Scott, 1994).
Breeding habitat - Marbled salamanders are one of two species of Ambystoma that breed on land (Petranka, 1998), and they are the only Ambystoma species that exhibit parental care (Nussbaum, 1985, 1987). Due to the terrestrial reproductive habits of Marbled Salamanders, breeding is restricted to fish-free wetlands with seasonally fluctuating water levels that include upland hardwood “swamp forests” (King, 1935), bottomland hardwood pools (Viosca, 1924a; Petranka and Petranka, 1981a,b), quarries (Graham, 1971), vernal ponds (Doty, 1978), Carolina bays (Jackson et al., 1989; Gibbons and Semlitsch, 1991), and floodplain pools (Petranka, 1990). Females remain with eggs (Noble and Brady, 1933) for varied lengths of time (Petranka, 1998); they may leave before eggs are inundated (McAtee, 1933; Jackson et al., 1989; Petranka, 1990). Nest brooding appears to enhance embryonic survival (Petranka and Petranka, 1981b; Jackson et al., 1989), although the mechanism is unknown. Opinions differ on whether there is an energetic cost to brooding by females (Kaplan and Crump, 1978; versus D.E.S., unpublished data). Occasionally nests are communal (Graham, 1971; Petranka, 1990), especially if cover items are scarce (Palis, 1996b).
Egg deposition sites - Breeding sites are generally the dried beds of temporary ponds, the margins of reduced ponds, or dry floodplain pools. Female Marbled Salamanders construct nests and lay eggs under virtually any cover in situations where the nest is likely to be flooded by subsequent winter rains (Noble and Brady, 1933). Eggs are laid on the edges of pools (Dunn, 1917b) and in dry basins under vegetation (Jackson et al., 1989), logs (Bishop, 1924; Doody, 1996), and leaf debris (Deckert, 1916; Petranka and Petranka, 1981b). Eggs are laid occasionally on non-soil substrate (Brimley, 1920a). Nest site selection by females is influenced by microsite elevation within the pond bed, site hydrologic regime, cover availability, and soil moisture (Petranka and Petranka, 1981a,b; Jackson et al., 1989; Figiel and Semlitsch, 1995; Wojnowski, 2000; but see also Marangio and Anderson, 1977). Females actively excavate oblong to ovoid-shaped depressions (King, 1935; Petranka and Petranka, 1981b).
Clutch size - Of the three reproductive modes of salamanders outlined by Salthe (1969), Marbled Salamanders have an atypical type I mode (Salthe and Mecham, 1974; Kaplan and Salthe, 1979). Clutch size ranges from approximately 30 to < 200 eggs (see Petranka, 1998) and usually is positively correlated with female body size (Kaplan and Salthe, 1979; Walls and Altig, 1986; Petranka, 1990; Scott and Fore, 1995), although not always (Kaplan and Salthe, 1979)—larger females may have larger eggs (Kaplan and Salthe, 1979).
Altig & McDiarmid 2015 - Classification and Description:
- Eastern Other - Oviposited as independent arrangements other than arrays or probable arrays that are difficult to recognize because of the small number of eggs or non-uniform substrate; most often in nonflowing water.
- Arrangement 1
- Sub-arrangement A - Eggs grouped singles, weakly adherent eggs, guarded by parent at semi terrestrial sites before autumnal rains fill pools; jelly layers tough and resilient.
- Arrangement 1
Compared to other species of Ambystoma, females in some populations of Marbled Salamanders may have fewer, larger eggs than would be expected for an animal of their size (Kaplan and Salthe, 1979; D.E.S., unpublished data) for a different opinion see Nussbaum, 1985, 1987). For example, comparing female Marbled Salamanders and mole salamanders (A. talpoideum) of equal body size, Marbled Salamanders have 3–4 times fewer eggs, but each egg is 3–4 times larger with 3–4 times higher lipid amounts (Komoroski, 1996; D.E.S., unpublished data). Mean egg diameter is greater in Marbled Salamanders than in flatwoods salamanders (A. cingulatum; 2.8 vs. 2.3. mm; Anderson and Williamson, 1976). Mean egg dry mass is greater in Marbled Salamanders than in either mole salamanders or spotted salamanders (A. maculatum; Komoroski, 1996). The caloric content (cal/mg dry mass) of Marbled Salamander eggs is greater than the energy content of spotted salamanders and tiger salamander (A. tigrinum) eggs (Kaplan, 1980b). Relatively few, large eggs with lipid stores in excess of the amount needed for embryogenesis probably reflects a response to the terrestrial breeding habits of Marbled Salamanders and the extreme variability and unpredictability in the timing of hatching (i.e., the duration of the egg stage). Substantial (15–30%) variation in egg diameter occurs within and among populations (Kaplan, 1980a). Egg size is positively correlated with hatchling size and early larval size (Kaplan, 1980a).
In spite of terrestrial egg laying, egg structure in Marbled Salamanders is similar to aquatic Ambystoma species (Salthe, 1963). Egg development is temperature-dependent (Noble and Brady, 1933); development (at similar temperatures) is slower than for some other ambystomatids (Moore, 1939). The prospective neural tissue of Marbled Salamanders has a lower density and higher water-holding capacity than the embryonic tissue of aquatic breeders such as spotted salamanders and tiger salamanders (M.G. Brown, 1942). Embryos develop to a hatching stage, but do not hatch until stimulated by hypoxia when the nest is flooded (Petranka et al., 1982). Some eggs may remain viable several months after oviposition (Noble and Brady, 1933), but often eggs laid in October will die by December if the nest has not been flooded (McAtee, 1933). An embryo’s lipid reserves in excess of reserves required for embryogenesis constrain the maximum time an embryo can survive in the egg (unpublished data). Hatching under natural conditions may occur at a wide range of developmental stages (Noble and Brady, 1933; Graham, 1971), probably due to the hydration state during development and the timing of nest inundation (Noble and Brady, 1933; S. Dooty, personal communication). Larvae will hatch from early stages (10.5 mm), with much yolk and little swimming ability, to late stages (20 mm; Brimley, 1920a; McAtee, 1933; Noble and Brady, 1933). By 18 mm the balancers are usually lost whether or not the egg has hatched (Brandon, 1961). Embryo mortality can be high due to freezing, dehydration, predation, or fungus (Stenhouse, 1987; Jackson et al., 1989) and is dependent on the timing and extent of pond-filling.
Timing of hatching varies among sites and years (Petranka and Petranka, 1980). Within a site, pond-filling may be incremental or sudden; gradual pond-filling may result in staggered hatching of eggs and substantial size variation of larvae within a pond (Smith, 1988).
Length of larval stage - Hatchling densities average as high as 47 larvae/m2 (Smith, 1988). Catastrophic larval mortality may result from winter kill due to extreme cold (Heyer, 1979; Cortwright and Nelson, 1990), incomplete pond filling and subsequent drying (King, 1935; Petranka and Petranka, 1981a), and early pond drying (Pechmann et al., 1991).
Larval growth rates of Marbled Salamanders vary considerably (see Doody, 1996; table 1), depending upon their density, food levels, and temperature. Growth rates are comparable to spotted salamanders (Walls and Altig, 1986), but comparisons to mole salamanders differ (Keen et al., 1984; Walls and Altig, 1986). Although larval growth is temperature dependent (Stewart, 1956), temperature effects may not be as pronounced as in some other Ambystoma species (Keen et al., 1984). Food level, temperature, hatching time, and larval density affect traits of newly metamorphosed animals (Stewart, 1956; Boone et al., 2002). Early hatching larvae are larger at metamorphosis, have higher survival, and metamorphose earlier than late-hatching larvae (Boone et al., 2002). Higher food levels and warmer temperature promote earlier metamorphosis; increased prey density promotes larger size at metamorphosis (Stewart, 1956). Early-hatching larvae are larger at metamorphosis, have higher survival, and metamorphose earlier than late-hatching larvae (Boone et al., 2002). Intraspecific larval density affects larval growth and a suite of larval traits (Stenhouse et al., 1983; Stenhouse, 1985b; Smith, 1988, 1990; Petranka, 1989c; Scott, 1990). At high densities, larvae have slower growth, smaller size at metamorphosis, and reduced survival (Petranka, 1989c); they may also have longer larval periods (Scott, 1990). As ponds dry, larval densities may become extremely high (e.g., 237 m2; Smith, 1988).
Larvae Food - Larvae eat primarily macrozooplankton, beginning with copepod nauplii in hatchlings (Petranka and Petranka, 1980). Ostracod, cladoceran, and copepod zooplankton feed larger larvae. Other invertebrate prey include chironomids, amphipods, chaoborids, and isopods (Petranka and Petranka, 1980; Branch and Altig, 1981).
Cover - Larvae may remain mostly hidden on the pond bottom during the day and move into the water column at night (Anderson and Graham, 1967; Petranka and Petranka, 1980; Branch and Altig, 1981). Both the limited diurnal movements and increased nocturnal activity may serve to enhance feeding and avoid vertebrate predation (Hassinger et al., 1970; Branch and Altig, 1981), although larvae floating in the water column at night did not capture more prey than those feeding on benthos during daylight (Petranka and Petranka, 1980). Movements of larvae into the water column are associated with decreased light intensity (Hassinger and Anderson, 1970) as well as vertical stratification of some prey species (Anderson and Graham, 1967; Petranka and Petranka, 1980). Larval activity may also vary seasonally; larvae remain near the bottom of the water column early in season and utilize the entire column for feeding on zooplankton as season progresses (Hassinger et al., 1970). Larvae nearing metamorphosis remain near the bottom at night (Petranka and Petranka, 1980).
Larval polymorphisms - None reported, although behavior differences are known. Laboratory assays have demonstrated two divergent aspects of kin recognition. In some contexts, kin recognition may reduce aggression and cannibalism among siblings in larval Marbled Salamanders (Walls and Roudebush, 1991); whereas in other contexts, large larvae may eat siblings preferentially (Walls and Blaustein, 1995). Hokit et al. (1996) further demonstrated that kin discrimination is context dependent. Under more natural conditions, kinship effects on larval performance did not occur (Walls and Blaustein, 1994).
Features of metamorphosis - As noted above, at high larval densities individual larvae have slower growth, a smaller size at metamorphosis, and reduced survival (Petranka, 1989c); they may also have longer larval periods (Scott, 1990). In field experiments, environmental conditions (i.e., larval density) accounted for more of the variation in body size at metamorphosis than did an individual’s level of multilocus genetic heterozygosity (Chazal et al., 1996). In an artificial pond study, more heterozygous individuals had shorter larval periods than less heterozygous larvae (Krenz, 1995). Juvenile recruitment into the terrestrial population may vary dramatically among sites and years (Stenhouse, 1984, 1987; Pechmann et al., 1991; Taylor and Scott, 1997), which probably reflects broad variation in abiotic (e.g., hydroperiod) and biotic (e.g., productivity, competition, and predation) conditions (Petranka, 1989c; Semlitsch et al., 1996). Effects initiated by aquatic conditions persist in terrestrial adults (Scott, 1994). Timing of metamorphosis may vary geographically, although recently metamorphosed juveniles generally disperse from the breeding site in late spring. Metamorphosis occurred in June–July in Illinois (Smith, 1961); June in New York (Bishop, 1941b); late May to early June in Maryland (Worthington, 1968), New Jersey (Hassinger et al., 1970), and north Georgia (Martof, 1955); mid May in West Virginia (Green, 1955); mid April to May in North Carolina (Stewart, 1956; Smith, 1988); March–April in Alabama (Petranka and Petranka, 1980); and as early as mid March in Louisiana (Dundee and Rossman, 1989). Although Marbled Salamanders will metamorphose in response to pond drying, timing also appears to be triggered by intrinsic factors (Hassinger et al., 1970; Scott, 1994). Larvae that hatch 2–3 mo later than others will nonetheless metamorphose within a few weeks of early hatching larvae, but at a smaller body size (unpublished data). Stages of metamorphosis are described by Grant (1931)
Post-metamorphic migrations - Juveniles may not disperse far from the edge of wetlands (P.K. Williams, 1973) and therefore require intact terrestrial habitats surrounding the breeding sites (Semlitsch, 1998).
Neoteny - There are no reports of non-transforming Marbled Salamanders. Given the widespread distribution and numerous population studies on this species, it is unlikely that neotenic adults exist.
Juvenile Habitat - Same as adult habitat, although juveniles tend to occur under smaller cover objects (Parmelee, 1993). Juveniles retain the ability to discriminate their siblings, presumably by chemoreception, for ≥ 8 mo after metamorphosis (Walls, 1991). Juvenile marbled salamanders experienced low first-year survival (4.5%) in old field terrestrial enclosures when compared to survival in forest enclosures (45%; Rothermel, 2003). Juveniles > 1-yr old experienced near zero annual survivorship in old field enclosures compared to > 70% in forest enclosures (Rothermel, 2003).
Adult Habitat - Most reports of terrestrial habitats indicate that mature deciduous forests are preferred (Petranka, 1998). Mixed hardwood and pine stands (Smith, 1988; Pechmann et al., 1991), floodplains (Petranka, 1989c; Parmelee, 1993), and uplands (Smith, 1961) are also utilized. Of 15 radioactively tagged individuals released near a woodland pond in southern Indiana, 14 were tracked in hardwood forest, 1 in an old field (P.K. Williams, 1973). Microhabitats within the forest include under leaf litter and small mammal burrows (P.K. Williams, 1973; Douglas and Monroe, 1981). Salamanders do not actively dig their own burrows, but enlarge existing openings (Semlitsch, 1983a). Although generally described as woodland salamanders, Marbled Salamanders may also be tolerant of relatively dry conditions (Cagle, 1942; Smith, 1961; Mount, 1975; Dundee and Rossman, 1989) and can be found on rocky hillsides (Johnson, 1987). One laboratory experiment indicated a preference for relatively basic substrates (pH 7.7), although animals in the field were found on more acidic (pH 5.5) substrates (Mushinsky, 1975). Compared to other ambystomatids, Marbled Salamanders may use substantially drier habitat and tolerate higher substrate temperatures (Parmelee, 1993). Adults dispersed an average of 194 m from the wetland breeding site (P.K. Williams, 1973). Consequently, post-metamorphic individuals require intact terrestrial habitats surrounding the breeding sites (Semlitsch, 1998). Survivorship of Marbled Salamander adults and recently metamorphosed animals was low in 100 m2 enclosures in clearcuts compared to enclosures in adjacent forests (P. Niewiarowski and A. Chazal, personal communication).
Home Range Size - Williams (P.K., 1973) examined home range size for Marbled Salamanders (n = 8) by using radioactive wire tags. The spring and summer home range size varied from 1–225 m2, with a median of 14.5 m2. There was a tendency for home range size to increase as individuals were followed for longer periods. A laboratory study indicated that juvenile Marbled Salamanders tend to stay on their own marked substrate, which may be a mechanism to detect home areas by chemical cues (Smyers et al., 2001).
Territories - Although Martin et al. (1986) found no evidence for territoriality in Small-mouthed Salamanders (Ambystoma texanum), they suggested that territoriality might be expected in Marbled Salamanders. Individuals of some Ambystoma species may return to their summer home range from the previous year (Semlitsch, 1983b), and this may be true in marbled salamanders (P.K. Williams, 1973). The orientation behavior exhibited by Marbled Salamanders is a necessary precursor to territoriality, although territoriality itself has not been documented. In laboratory studies, “resident” individuals tend to bite conspecific “intruders;" however, animals housed together for long periods did not avoid each other (Ducey, 1989). Juvenile salamanders (< 8 mo old) are less aggressive to familiar “neighbors” than to unfamiliar “strangers,” especially among siblings (Walls, 1991). Adult Marbled Salamanders maintained on a low-food diet were more prone to bite an intruding salamander than those on a high-food diet (Ducey and Heuer, 1991), which may indicate that aggression functions to repel an intruder from an individual’s feeding area/burrow refuge. Marbled salamanders almost always occur alone under cover objects (Parmelee, 1993).
Aestivation/Avoiding Dessication - Marbled Salamanders likely undergo prolonged periods of summer inactivity, corresponding to periods of little or no rainfall. Despite reports that Marbled Salamanders can occur in unusually dry habitats (e.g., Bishop, 1943), there is no evidence that they differ from more aquatic species in terms of their water exchange with soil (Spight, 1967b). However, a laboratory study of water loss rate showed Marbled Salamanders lose water more slowly than the other species examined (which were all plethodontids) and were able to withstand dehydration ≤ 30% of initial body weight (Spight, 1968). Dehydrated salamanders incur substantial metabolic costs, however (Sherman and Stadlen, 1986). To minimize water loss, Marbled Salamanders likely avoid desiccating conditions; as soils dry in late summer, animals may retreat to deeper burrows (P.K. Williams, 1973). Rehydration rates were faster in Marbled Salamanders than in more aquatic species, and faster in severely dehydrated animals than in less dehydrated individuals (Spight, 1967a).
Heat stress - Larvae of Marbled Salamanders have less resistance to high temperatures (i.e., have a lower Critical Thermal Maximum, CTM) than either Small-mouthed Salamanders or spotted salamanders (Keen and Schroeder, 1975). Smaller adult salamanders reach their CTM faster than larger adults (Hutchison, 1961). The possible relationship between CTM in eggs, larvae, and adults, and geographic distribution or timing of breeding (sensu Gatz, 1971) is unknown.
Water stress - In general, post-metamorphic Marbled Salamanders do not appear to respond well to prolonged immersion in water (personal observation). Interestingly, the total oxygen uptake through pulmonary surfaces is relatively low (34%; Whitford and Hutchison, 1966b), although lung sacs, ridges, and vascularization are well developed in Marbled Salamanders (Czopek, 1962). Under anoxic conditions, larvae may exhibit anaerobic glycolysis (Weigmann and Altig, 1975). iii. Metabolic rate. Lunged salamanders, including Marbled Salamanders, increase levels of oxygen consumption with increasing body size (Whitford and Hutchison, 1967; Krenz, 1995). A daily cycle also occurs, with resting metabolic rate increasing by 50% during the early evening (Krenz, 1995). Metabolic rates increase by 119% following dehydration (Sherman and Stadlen, 1986). Resting metabolic rate is positively correlated with multi-locus heterozygosity (Krenz, 1995); more heterozygous females with higher metabolic demands allocated less energy to their clutches of eggs (Krenz, 1995).
Seasonal Migrations - Restricted to times of breeding (adults; see "Breeding migrations" above) and following metamorphosis (juvenile; see "Features of metamorphosis" above). Post-metamorphic dispersal is restricted to rainy nights. The period between metamorphosis and dispersal may be several weeks or more (depending on occurrence of nighttime rainfall) and is likely a period of high mortality for juveniles (personal observation).
Torpor (Hibernation) - In the north, post-reproductive adult Marbled Salamanders move ≤ 30 m from the breeding site (Douglas and Monroe, 1981), where they remain for the winter. Hibernation in the southern portions of their range is unknown.
Interspecific Associations/Exclusions - Due to the terrestrial breeding habits and early egg hatching, larval Marbled Salamanders are often much larger than other amphibian larvae (Worthington, 1968, 1969; Keen, 1975; Stenhouse et al., 1983; Walls and Altig, 1986; Smith, 1988; Scott, 1993). Larval Marbled Salamanders will feed on other amphibian eggs and larvae (Walters, 1975), including Ambystoma larvae. Where they co-occur, Marbled Salamanders eat smaller Spotted Salamander larvae (Stewart, 1956; Stenhouse et al., 1983; Stenhouse, 1985b), Small-mouthed Salamanders (Walters, 1975; Doody, 1996), Jefferson Salamanders (A. jeffersonianum; Cortwright, 1988), tiger salamanders (Stine et al., 1954), and mole salamanders (Walls, 1995). Spotted Salamander larvae may be more susceptible than mole salamander larvae to this predation due to increased use of refugia by Mole Salamanders (Walls, 1995). The size of Marbled Salamander larvae at the time when other Ambystoma eggs are hatching varies among ponds and years by 30–40% (Stenhouse et al., 1983). Consequently, although larval Marbled Salamanders are often predators, they may also be competitors (Wilbur, 1984; Stenhouse, 1985b; Cortwright, 1988; Semlitsch et al., 1996). Predation by Marbled Salamander larvae may substantially affect community dynamics (Cortwright and Nelson, 1990; Morin, 1995; Boone et al., 2002). Juvenile Marbled Salamanders that were tested under laboratory conditions with conspecifics and with juvenile Mole Salamanders did not show any overt aggression, perhaps indicating that such behavioral interactions are not important for juveniles (Smyers et al., 2001). Additional experiments with juvenile Spotted Salamanders indicated that juvenile Marbled Salamanders may defend burrow space by excluding heterospecific salamanders (Smyers et al., 2002).
Age/Size at Reproductive Maturity - Age and size at reproductive maturity are traits that vary and are highly dependent on size at metamorphosis (Scott, 1994), which in turn is influenced by intraspecific larval densities and the timing of pond drying (Petranka, 1989c; Scott, 1990). Males tend to mature at an earlier age than females (Scott, 1994; Pechmann, 1995); average age at first reproduction for males is 2.5–3.1 yr (Scott, 1994) to 3.3 yr (Pechmann, 1995), and for females, 2.8–3.4 yr (Scott, 1994) to 4.0 yr (Pechmann, 1995). The range of age at first reproduction for both sexes is 1–7 yr. Mean size at first reproduction is approximately 53–60 mm SVL for both sexes (Scott, 1994; Pechmann, 1995); the minimum size at first reproduction may be smaller for males (~42.0 mm) than for females (~45.0 mm).
Longevity - Survival to first reproduction can be low and is influenced by size at metamorphosis. Variation in body size at metamorphosis is coupled with variation in lipid stores (ranging from 2–16.5% of dry mass; unpublished data). Small, lean animals may suffer the highest mortality immediately following metamorphosis (Scott, 1994). Survival from metamorphosis to first reproduction ranges from 3–60% (Scott, 1994; Pechmann, 1994, 1995). Males may exhibit higher survivorship than females due to their earlier age at first reproduction (Scott, 1994). Maximum lifespan in the field appears to be 8–10 yr (Graham, 1971; Taylor and Scott, 1997). N. Feeding Behavior. Stomach contents of juveniles and adults include millipedes, centipedes, spiders, insects, and snails (Dundee and Rossman, 1989); arthropods, annelids, and mollusks (Smith, 1961).
Eggs - Eggs may be preyed upon by beetles, salamanders, frogs (Noble and Brady, 1933), and possibly a millipede species (Uroblaniulus jerseyi; Mitchell et al., 1996a).
Larvae - Larvae are palatable to fishes (Kats et al., 1988), but usually do not inhabit ponds where fish occur. Larval Marbled Salamanders are prey for numerous species, especially invertebrates including dragonfly naiads (Odonata), spiders (Arachnida), dytiscid beetle larvae and adults (Coleoptera), and giant water bugs (Bellostomatidae). Larval survivorship decreased from 60 to 70% to < 20% when hatchlings inhabited experimental enclosures in a wetland replete with invertebrate predators (unpublished data); survivorship decreased to zero in a year when chain pickerel (Esox niger) colonized the wetland. Adult Eastern Newts (Notophthalmus viridescens) and paedomorphic Mole Salamanders also feed on larval Marbled Salamanders. Cannibalism may occur (Walls and Roudebush, 1991) when incremental pond-filling staggers dates of hatching and increases size variation (Smith, 1990). Wading birds and kingfishers (Megacerle alcyon) are also likely predators (personal observations).
Juveniles and adults - Raccoons (Procyon lotor), opossums (Didelphis virginiana), skunks (Mustelidae), and shrews (Soricidae) are known to kill adult Marbled Salamanders (DiGiovanni and Brodie, 1981; Petranka, 1998). Often the tails are not eaten (personal observation). Newly metamorphosed animals may be susceptible to mammalian predators as well as some snakes; one Banded Watersnake (Nerodia fasciata) had eaten 34 recently metamorphosed Marbled Salamanders (unpublished data). Liner (1954) reported ingestion of two recently metamorphosed Marbled Salamanders by a Western Ribbonsnake (Thamnophis proximus).
Eggs - Protection of eggs from predators is possibly one function of nest-brooding by females (Petranka, 1990).
Larvae - Limited diurnal movements and hiding in benthic debris may reduce predation (Hassinger et al., 1970; Petranka and Petranka, 1980; but see Marangio, 1975, for report of positive phototaxis in small larvae). Hatchlings and small larvae may use the sun as a cue to orient toward deep water (Tomson and Ferguson, 1972). Larvae do not change behavior (i.e., increase use of refugia) in the presence of fishes (Kats et al., 1988).
Adults - Animals under attack by short-tailed shrews (Blarina brevicauda) exhibit tail lashing, body coiling, and head-butting behaviors, and/or may become immobile (Brodie, 1977). Such behaviors may draw the attacks toward the tail, which has concentrations of granular glands on dorsum that produce noxious secretions. Adults are unpalatable to common ribbon snakes (Thamnophis s. sauritus; T. Mills, personal communication). Secretions generally confer protection from a single attack by shrews (Brodie et al., 1979). Secretions in marbled salamanders are reduced after multiple attacks by shrews, resulting in increased vulnerability (DiGiovanni and Brodie, 1981).
Diseases - An aquatic fungus (Saprolegnia sp.) may develop on the injured portions, especially limbs, of bitten larvae and may be lethal (Petranka, 1989c). Marbled salamanders have been used in toxicological tests of hydrazine compounds (Slonim, 1986), beryllium sulfate (Slonim and Ray, 1975), pesticides (Hall and Swineford, 1981), and motor oil (Lefcourt et al., 1997).
Parasites - Rankin (1937) reported the following parasites from Marbled Salamander larvae in North Carolina: Protozoa—Cryptobia borreli, Eutrichomastix batrachorum, Hexamitus intestinalis, Prowazekella longifilis, Tritrichomonas augusta; Trematoda—Diplostomulum ambystomae; Acanthocephala—Acanthocephalus acutulus. Rankin (1933) reported the following parasites from Marbled Salamander adults in the same populations: Protozoa—Cryptobia borreli, Cytamoeba bacterifera, Eimeria ranarum, Eutrichomastix batrachorum, Haptophyra michiganensis, Hexamastix batrachorum, Hexamitus intestinalis, Prowazekella longifilis, Tritrichomonas augusta; Trematoda—Brachycoelium hospitale, Diplostomulum ambystomae; Gorgoderina bilobata, Megalodiscus temperatus, Plagitura sp.; Nematoda—Capillaria inequalis, Cosmocercoides dukae, Filaria sp., spirurid cysts; Acarina—Hannemania dunni.
The trematode Brachycoelium ambystomae was reported from Marbled Salamanders by Couch (1966), and an unidentified immature trematode by Malewitz (1956). The gall bladder myxosporean (Myxidium serotinum) has been reported in Marbled Salamanders in Arkansas and Texas (McAllister and Trauth, 1995).
Conservation - Marbled salamanders are listed as Threatened in Massachusetts and Michigan, and Protected in New Jersey (Levell, 1997). In each of these states, permits are required for any activity involving Marbled Salamanders.
Given the reliance of Marbled Salamanders on small isolated seasonal wetlands and intact forested floodplain habitats, their abundance presumably has declined as wetland habitats have been destroyed (Petranka, 1998). Small isolated wetlands are the most valuable wetlands for maintaining amphibian biodiversity, but it is precisely these wetlands that are unprotected by current wetlands regulations and that are most “at risk” (Semlitsch and Bodie, 1998). Further loss of small wetlands such as Carolina bays will likely be accelerated by the U.S. Supreme Court’s SWANCC decision in January 2001 (Sharitz, 2003), unless individual states pass legislation that protects small isolated wetlands. As isolated wetland habitats disappear and remaining wetlands become increasingly separated, the cumulative impact on amphibian populations such as Marbled Salamanders will likely be substantial and perhaps non-linear, as elimination of remaining wetlands results in proportionally larger and larger effects on pond-breeding amphibian populations. For conservation efforts to succeed it will also be critical that the wetland ecosystem be viewed not solely as the wetland itself, but also the adjacent terrestrial habitat that is essential to the persistence of pond-breeding amphibians (Scott, 1999; Gibbons, 2003).
References for Life History
- Altig, Ronald & McDiarmid, Roy W. 2015. Handbook of Larval Amphibians of the United States and Canada. Cornell University Press, Ithaca, NY. 341 pages.
- AmphibiaWeb. 2020. University of California, Berkeley, CA, USA.
- Bishop, S.C., 1941, The salamanders of New York, New York State Mus. Bull., Vol. 324, pg. 1-365
- Graham, R.E., 1971, Environmental effects on deme structure, dynamics, and breeding strategy of Ambystoma opacum (Amphibia: Ambystomatidae), with an hypothesis of the probable origin of the marbled salamander life-style, Ph.D. Diss., Rutgers Univ., New Brunswick, N.J., UNPB
- Hassinger, D.D., Anderson, J.D., Dalrymple, G.H., 1970, The early life history and ecology of Ambystoma tigrinum and Ambystoma opacum in New Jersey, Am. Midl. Nat., Vol. 84, pg. 474-495
- King, W., 1935, Ecological observation on Ambystoma opacum, Ohio J. Sci., Vol. 35, pg. 4-15
- Martof, B.S., 1955, Observations on the life history and ecology of the amphibians of the Athens area, Georgia, Copeia, Vol. 1955, pg. 166-170
- Minton, S.A., 1972, Amphibians and Reptiles of Indiana, Indiana Academy of Science Monograph, Vol. 3, 346 pgs., Indiana Academy of Science, Indianapolis
- Petranka, J.W., Petranka, J.G., 1981, On the evolution of nest site selection in the marbled salamander, Ambystoma opacum, Copeia, Vol. 1981, pg. 387-391
- Barbour, R.W., 1971, Amphibians and reptiles of Kentucky, 334 pgs., Univ. of Kentucky Press, Lexington, KY
- Bishop, S.C., 1943, Handbook of Salamanders, 555 pgs., Comstock Publ. Co., New York, NY
Photos:
*Click on a thumbnail for a larger version.


402.JPG)
.JPG)






Verified County/City Occurrence
Accomack
Albemarle
Alleghany
Amelia
Amherst
Appomattox
Arlington
Augusta
Bath
Bedford
Botetourt
Brunswick
Buckingham
Campbell
Caroline
Charles City
Charlotte
Chesterfield
Craig
Culpeper
Cumberland
Dinwiddie
Fairfax
Fauquier
Floyd
Fluvanna
Franklin
Gloucester
Goochland
Greensville
Halifax
Hanover
Henrico
Henry
Isle of Wight
James City
King and Queen
King George
King William
Lancaster
Loudoun
Louisa
Lunenburg
Madison
Mathews
Mecklenburg
Middlesex
Montgomery
Nelson
New Kent
Northampton
Northumberland
Nottoway
Orange
Page
Patrick
Pittsylvania
Powhatan
Prince Edward
Prince George
Prince William
Pulaski
Rappahannock
Richmond
Roanoke
Rockbridge
Rockingham
Scott
Shenandoah
Southampton
Spotsylvania
Stafford
Surry
Sussex
Westmoreland
Wise
York
CITIES
Chesapeake
Hampton
Newport News
Poquoson
Richmond
Suffolk
Virginia Beach
Williamsburg
Verified in 77 counties and 8 cities.
U.S. Range

Box Turtle Reporting
