Spotted Salamander
Ambystoma maculatum
Common Name: |
Spotted Salamander |
Scientific Name: |
Ambystoma maculatum |
Etymology: |
|
Genus: |
Amby is Greek for "a cup", stoma is Greek for "a mouth" |
Species: |
maculatum is Latin for "spot" or "spotted" |
Average Length: |
4.4 - 7.8 in. (11.2 - 19.7 cm) |
Virginia Record Length: |
|
Record length: |
9.8 in. (24.8 cm) |
Physical Description - This species possesses a dorso-lateral series of rounded yellow or orange spots. The sides are dark gray or slate and the belly pale slate without spots or blotches. There are usually 12 costal grooves and the tongue has plicae radiating from the posterior field. The length of this species is up to 206 mm. The males are slightly slimmer than the females and in the breeding season they may be recognized by the protuberant vent. The females at this season have the ventral surfaces paler.
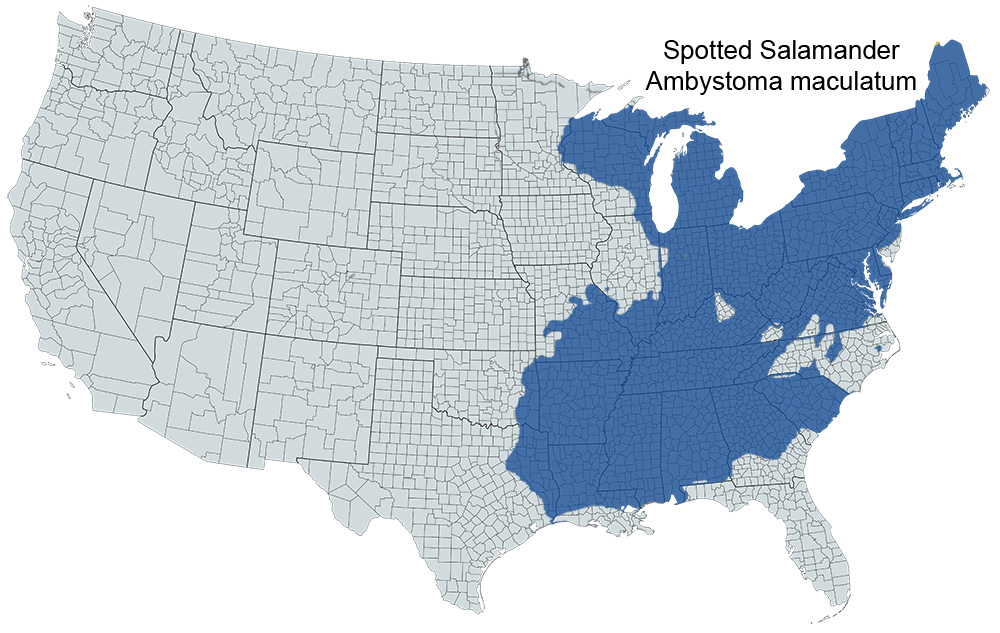
Historical versus Current Distribution: Spotted salamanders (Ambystoma maculatum) are distributed throughout the interior and Atlantic Coastal Plains of the eastern United States, primarily in hardwood and mixed coniferous-deciduous forest habitats (Bishop, 1943; Pope, 1944; Anderson, 1967b; Upchurch, 1971; Lang, 1972; Schuette, 1980; Kats et al., 1988; Lovich and Fisher, 1988; Beane and Gaul, 1991; C. Phillips, 1991a,b, 1992, 1994; Schwaner and Anderson, 1991; E.E. Brown, 1992; Petranka, 1998). This species prefers lowland forests; they are occasionally found in more open habitats such as meadows, but usually near forest edges. Although not typically a montane species, spotted salamanders can occur at higher elevations when suitable breeding sites are available (Thompson and Gates, 1982). With the exception of a few isolated populations, spotted salamanders are also absent from the Coastal Plain of North Carolina, as well as southeastern portions of Virginia, Maryland, Delaware, and New Jersey. No specimens have been recorded from the Florida Peninsula or the Coastal Plain of the Florida Panhandle to southern Georgia.
There is no evidence to suggest that current and historical distributions differ; however, range reductions undoubtedly have occurred, and continue to occur, due to habitat alteration. Despite these changes, new county records (e.g. Brinkman et al., 2001; Cochran and Cochran, 2001; Scott et al., 2001) documenting the geographic distribution of spotted salamanders have been published in recent years. Moreover, two new county records extend the range of this species into Minnesota (C. Hall, in review), indicating that the extent of the range is still being identified.
Smith (1961) mentioned geographic variation between populations in northeast and southern Illinois in the average sizes of adults, number of teeth, number of dorsal spots, costal groove counts, and contrast between head spot and body spot color. Likewise, DuShane and Hutchinson (1944) raised larvae from independent populations in a common garden experiment and demonstrated significant differences between these populations in developmental rates and size at metamorphosis. This variation has led some to suggest the presence of two distinct subspecies of spotted salamanders (Smith, 1961).
There is indirect evidence that ranges may have been reduced substantially during the Pleistocene glacial cycles, causing geographic discontinuity and resulting in the genetic differentiation between two highly differentiated lineages (C. Phillips, 1994; Zamudio and Savage, 2003). Post-glacial recolonization resulted in the expansion and recontact of these two lineages in at least three major regions: (1) the southern Interior Lowlands of Missouri and Illinois; (2) the Interior Lowlands of Ontario, Canada; and (3) from the southern Appalachian Highlands to the coast of South Carolina (Zamudio and Savage, 2003). Three fossil vertebral elements assigned to spotted salamanders date from the late Pleistocene and were collected in a cave near St. Louis, Missouri (Holman, 1965b), near the western extent of the species' current range. A second fossil form (A. minshalli) has been proposed as ancestral to A. maculatum (Holman, 1975) and is clearly a member of the maculatum group. These fossils, collected in Nebraska and Kansas (Estes and Tihen, 1963; Tihen and Chantell, 1963), date back to the Upper Miocene, suggesting that this group was well differentiated before Plio-Pleistocene climatic changes. The presence of fossils in these regions indicates that the historical range of spotted salamanders coincides in part with the contemporary range and that members of the spotted salamander lineage once extended farther west into the Great Plains.
Historical versus Current Abundance Although there are no quantitative assessments of declines in abundance for spotted salamanders, their habitat requirements suggest that, as with other pond-breeding species, they are adversely affected by deforestation and wetland destruction (Petranka, 1998). Habitat loss, acidification, metal concentrations, environmental contaminants, and fish introductions are certainly factors that could contribute to declines of spotted salamanders, and some have been shown to negatively affect this species (Pough, 1976; Clark, 1986b; Blem and Blem, 1989, 1991; Sadinski and Dunson, 1992; Brodman, 1993; Rowe and Dunson, 1993; but see Cook, 1983). Acid rain has received particular attention in the Northeast as a factor reducing survivorship (e.g., Portnoy, 1990). Future spotted salamander declines are possible because these animals have not evolved mechanisms to tolerate acidic conditions or introduced fishes, suggesting they will respond negatively to these threats (Tome and Pough, 1982; Clark, 1986b; Sexton et al., 1994; see also Petranka, 1998). Because overall range reductions caused by changing landscape features coincide with human land use, we expect that current abundances are lower than historical numbers.
Breeding: Reproduction is aquatic.
Breeding migrations: Many life history aspects of spotted salamanders vary tremendously across their geographic range, most likely due to large differences in environmental conditions. The pattern of variation is complex and does not seem to be determined exclusively by latitude or geographic locality. Throughout their range, individuals migrate from terrestrial overwintering sites into seasonally available aquatic habitats to breed. Although the vast majority of individuals are reproductively active, a small percentage of non-reproductive individuals also migrate (Shoop, 1967). Why these non-reproductive animals migrate is not known. Breeding individuals are easily identified by body size and characteristics of the vent: males have a conspicuously swollen vent covered with rough papillae; females have a rounded, smooth vent, and are generally larger in body size (Wilson, 1976; Sexton et al., 1986). Finkler et al. (2003) identified sex-related differences in terrestrial and aquatic locomotor performance in breeding spotted salamanders.
Males typically arrive earlier, anywhere from 1–6 d, than females at ponds (Bishop, 1941b; Hillis, 1977; Douglas, 1979; Sexton et al., 1986). This appears to be due to a differential response between the sexes to temperature cues for the onset of migration (Sexton et al., 1990) or to locomotor performance (Finkler et al., 2003). Migrations take place at night during or following early spring rains correlated with increasing air temperatures; in more northern regions, migrations follow periods of increased temperature and heavy snowmelt or rain (Wright, 1908; Wright and Allen, 1909; Blanchard, 1930; Bishop, 1941b, 1943; Baldauf, 1952; Whitford and Vinegar, 1966; Wilson, 1976; Hillis, 1977). The exact role of each of these ecological features in the onset of migrations is still unclear. A detailed study by Sexton et al. (1990) showed a threshold system of rainfall and temperature cues and a correlation of immigration with soil temperatures (4.5 ˚C, 30 cm below the surface) when the thermal profile was reversed.
The onset of breeding varies greatly across their range and can occur as early as December in southern populations or as late as April at higher latitudes (Brimley, 1921a; Blanchard, 1930; Dempster, 1930; King, 1939; Pawling, 1939; Welter and Carr, 1939; Gray, 1941; Baldauf, 1952; Hardy, 1952; Moulton, 1954b; Peckham and Dineen, 1954; Green, 1955; Seibert and Brandon, 1960; Whitford and Vinegar, 1966; Worthington, 1968, 1969; Minton, 1972, 2001; Keen, 1975; Mount, 1975; Harris, 1980; Walls and Altig, 1986; Talentino and Landre, 1991; Pinder and Friet, 1994; Brodman, 1995). There is a rough latitudinal pattern in the degree of breeding synchrony within populations: in more southern populations the breeding season may last over 2 mo, usually with 2–3 major bouts of activity following heavy rains (Peckham and Dineen, 1954; Mount, 1975; Harris, 1980; Sexton et al., 1986); breeding in northern regions is more synchronized and can be restricted to a few days (2–5 d; Wilson, 1976; Talentino and Landre, 1991; Brodman, 1995; Petranka, 1998). In a Missouri population, Sexton et al. (1986) estimated that the breeding season lasts 45 d for males, 37 d for females. In North Carolina, Stenhouse (1985a) reported 56 and 73 d in consecutive years. Husting (1965) recorded breeding seasons of 9–29 d over 5 yr in Michigan; in an Indiana population, breeding averaged 25 d (Peckham and Dineen, 1954). Over 4 yr, Wilson (1976) recorded breeding seasons that averaged 18.5 d. By contrast, breeding seasons in Ohio lasted only 3–5 d (Brodman, 1995); 2–3 d in Massachusetts (Talentino and Landre, 1991). During the breeding season both sexes lose weight. In Missouri, Sexton et al. (1986) reported average weight losses of 13% and 38%, for males and females, respectively; whereas in Massachusetts, Windmiller (1996) found values of 4.5% and 24.1%.
Return rates of adults to breeding ponds suggest that while individuals in some populations breed annually, others may skip years; however, mark–recapture studies may underestimate the number of animals breeding in consecutive years because they do not account fully for dispersal of adults to other sites. Thus it is unknown whether adults are capable of sustained annual breeding or if the sexes differ in that regard. High annual return rates were recorded at a Rhode Island breeding pond (89.5%; Whitford and Vinegar, 1966). Douglas and Monroe (1981) reported six of eight (75%) tagged individuals returning to a Kentucky breeding pond in two consecutive years; however, these samples sizes were relatively small. In Missouri, 38% of marked females and 30% of marked males returned to breed the following year (Phillips and Sexton, 1989). Less consistent breeding patterns were found in a Michigan population, where annual return rates ranged from 18–55% over the course of five years (Husting, 1965). In New York, Wilson (1976) recaptured a total of only 12 males and 1 female in the four years of his study, representing 8.7% of males and 0.7% of females. These longer-term studies suggest that there may be large temporal as well as inter-population differences in breeding patterns. Numerous studies have investigated migratory orientation of spotted salamanders during the breeding season. Adults usually exit ponds near where they entered, suggesting that they use the same migratory route between terrestrial home ranges and breeding sites (Shoop, 1965a, 1968; Wilson, 1976; Douglas and Monroe, 1981; Kleeberger and Werner, 1983; Stenhouse, 1985a; Sexton et al., 1986; Phillips and Sexton, 1989; see also Petranka, 1998). In addition, the order of male arrival was correlated between years in one population in North Carolina (Stenhouse, 1985a), suggesting that males may be returning to the same home ranges after breeding. Downs (1989d) observed some adults returning to the same home burrow after emigrating from breeding ponds in Ohio.
Post-breeding adults move on average 115.7 m from the pond edge to their terrestrial home ranges (range, 0–249 m; average of adult movement distances reported from six studies; Wacasey, 1961; P.K. Williams, 1973; Douglas and Monroe, 1981; Kleeberger and Werner, 1983; Madison, 1997). Semlitsch (1998) reviewed adult movement patterns in six species of ambystomatids (including spotted salamanders) and concluded that terrestrial buffer zones of breeding ponds would have to extend 164.3 m from the edge to encompass movements of 95% of the adults in a population.
Mark–recapture studies and displacement experiments suggest that two cues may be important in migratory orientation. Olfaction was suggested as an orientation mechanism in a laboratory choice experiment where adult salamanders preferred substrate saturated with their home pond odors (McGregor and Teska, 1989). Shoop (1968) and Whitford and Vinegar (1966) displaced adult individuals 500 m and 128 m from their home ponds. Most individuals successfully oriented back to their home ponds; a higher percentage of those released in and adjacent to a seepage returned to their home pond, suggesting they may be using rheotaxic orientation in addition to olfaction (Whitford and Vinegar, 1966). Rheotropism was also suggested in individuals migrating through runoff in two Michigan ponds following heavy rains (Finneran, 1951). These data are suggestive but not conclusive and underscore the need for a rigorously controlled experimental test of orientation mechanisms.
Breeding habitat: In addition to ephemeral, fishless wetlands, spotted salamanders will breed in roadside ditches and tire ruts in dirt roads, artificial ponds, floodplain wetlands, and marshes. Although they typically use ephemeral habitats, breeding has been documented in permanent ponds containing fishes (Husting, 1965; Harris, 1984; Figiel and Semlitsch, 1990; see also Petranka, 1998), as well as in a fish-free lake in New York (Bahret, 1996). Populations in the Atlantic Coastal Plain sometimes breed in stream backwaters containing predatory fishes; larvae typically will not survive in these habitats unless there is sufficient submergent vegetation (Husting, 1965; Harris, 1984; Figiel and Semlitsch, 1990).
In breeding ponds, adults aggregate in large polyandrous groups for courtship and mating, referred to as a nuptial dance or “liebesspiel” (O'Donnell, 1937). In the presence of females, courting males deposit spermatophores in an area of approximately 1 m2 on the pond bottom. Spermatophores are usually attached to leaves, twigs, and other submerged vegetation (Breder, 1927; Bishop, 1943; Petranka, 1998). Arnold (1976) showed in experimental field enclosures that males produce on average 40 spermatophores the first night when in the presence of a female (range, 10–81), and spermatophore production decreases rapidly on successive nights. A male vigorously courts a female by repeatedly nudging her with his snout, swinging his head back and forth along her dorsum, and lifting his head under her chin (Arnold, 1976). A sexually receptive female will also nudge males and search for spermatophores by stepping from side to side with her hindlimbs while moving forward. Sound production has been reported during lulls in courtship (Breder, 1927), although it is unknown whether there is any courting significance because sounds were emitted more often in the presence of same-sex individuals (Wyman and Thrall, 1972 ). Once a female finds a spermatophore, she orients towards it tactually, squats, arches the base of her tail and removes the sperm mass from the gelatinous base with her vent. Females pick up multiple sperm masses, and paternity analyses of experimental matings (Tennessen and Zamudio, 2003) and wild-collected clutches (E.M. Myers and K.R.Z., unpublished data) confirm that multiple paternity is common in this species and that eggs are frequently fertilized by sperm storage. Arrival time at breeding ponds has significant impact on male reproductive success. In experimental enclosures in the field, Tennessen and Zamudio (2003) enclosed mating trios (one female and two males) in control enclosures and both males were allowed to mate at the same time; in experimental enclosures one male was introduced in the mating chamber 2 hr before the second male. Paternity analyses of the resulting offspring indicate that early arrival at a mating aggregation is an important determinant of reproductive success. In control enclosures, the two males fathered similar proportions of offspring; in experimental enclosures, the early arriving male sired significantly more offspring than the late-arriving male (Tennessen and Zamudio, 2003).
Sex ratios of spotted salamanders are always male-biased in breeding ponds and vary from 1.5–3.5:1 (Peckham and Dineen, 1954; Husting, 1965; Whitford and Vinegar, 1966; Hillis, 1977; Stenhouse, 1985a; Sexton et al., 1986; Downs, 1989d; Flageole and Leclair, 1992). Differences in age at maturity, reproductive frequency, and survival may explain the strongly male-biased sex ratios (Wacasey, 1961; Flageole and Leclair, 1992). This ratio, combined with the short breeding season, produces extreme sexual competition among males. Sexual interference by spermatophore “capping” is a common behavior for males. When males encounter other spermatophores, they deposit a secondary spermatophore upon it, but it is unknown whether males can discriminate their own spermatophores from those of other males (Arnold, 1976). Spermatophores can be found in stacks 2–6 high, and the female presumably picks up the top-most sperm mass.
Egg deposition sites: Egg masses are either scattered individually in breeding ponds or deposited in aggregates of 50 clutches or more. Single eggs have been found on land near a pond (Brimley, 1921a; Smith, 1921). Reported mean numbers of eggs/mass are highly variable across regions, ranging from 58–155 (Bishop, 1941b; Seibert and Brandon, 1960; Woodward, 1982; Stangel, 1988; Downs, 1989d; Talentino and Landre, 1991). Egg masses are deposited in firm and compact masses on sticks, submergent vegetation, small branches, or directly on the pond bottom. The thick, firm jelly coat on eggs serves to protect them from desiccation (Nyman, 1987), physical disturbances, and predation (Cory and Manion, 1953; Ward and Sexton, 1981; Semlitsch, 1988).
Some females deposit eggs containing hydrophobic proteins in the outer jelly layer producing white or milky egg masses in which the embryos are not visible (Hardy and Lucas, 1991). These white and clear forms are present among individuals within populations, but there is no evidence of differences in egg viability or hatching size related to this polymorphism. However, Brodman (1995) demonstrated that white egg masses in an Ohio population contained significantly more eggs than clear egg clutches (103.5 ± 11.7 and 66.1 ± 4.3, respectively), suggesting the advantage may lie in higher fecundity. Metts (2001) also found that white egg masses contain more eggs than clear ones, but that clear egg masses have significantly better hatching success than white masses. The functional role or advantage of the white protein is still unclear (Ruth et al., 1993).
Clutch size: Females deposit 2–4 egg masses, each containing from 1–250 eggs (Bishop, 1941b; Shoop, 1974; Wilson, 1976; Petranka, 1998).
After egg deposition, chlamydomonad algae (Oophila amblystomatis) enter the inner jelly capsules of individual eggs and presumably aids in increased oxygen supply to developing embryos (Breder, 1927; Gilbert, 1942, 1944; Hutchison and Hammen, 1958; Hammen and Hutchison, 1962; Gatz, 1973a; Bachmann et al., 1986). In laboratory experiments, embryos with algal symbionts hatched sooner than clutches with reduced algal growth (Gilbert, 1942). Embryonic mortality is variable and can be high, ranging from 0–100% (Pough, 1976; Harris, 1980; Cook, 1983; Clark, 1986b; Stenhouse, 1987; Stangel, 1988; Ireland, 1989; Brodman, 1995). Eggs laid later in the breeding season had higher rates of survivorship in a North Carolina pond, suggesting that within-year timing of breeding can have important implications for individual fitness (Harris, 1980). Egg mortality is increased by low pH, predation, and freezing in shallow areas of ponds (Pough, 1976; Wilson, 1976; Ireland, 1989). Incubation times range from 8–60 d (Bishop, 1941b, 1943; Whitford and Vinegar, 1966; Worthington, 1968, 1969; Shoop, 1974; Keen, 1975; Freda, 1983; Sexton et al., 1986; Stangel, 1988; Nyman, 1991; Talentino and Landre, 1991), and hatchlings from a Mississippi pond measured 14.8 mm ± 1.46 (Walls and Altig, 1986). However, all developmental features are temperature-dependent (Voss, 1993a) and vary depending on the population and region (DuShane and Hutchinson, 1944).
Length of larval stage: Duration of the larval stage is highly variable and lasts anywhere from 6 wk to > 12 mo (Bishop, 1941b, 1943; Dundee, 1947; Worthington, 1968; Freda, 1983; Nyman, 1991; Talentino and Landre, 1991). Metamorphic patterns can vary within a population across years (Phillips, 1992), as well as among individuals within populations (Ireland, 1973). Most larvae transform from June–August (Petranka, 1998), but overwintering larvae have been found in spring-fed ponds (Bleakney, 1952; Whitford and Vinegar, 1966; Ireland, 1973; Hillis and Miller, 1976; Stangel, 1988; Phillips, 1992). Overwintering larvae in Missouri reach, on average, 69 mm TL and transform as late as May (Phillips, 1992). Larvae range in size from 27–75 mm TL at metamorphosis, and these lengths vary greatly across populations (Bishop, 1941b, 1943; Dundee, 1947; Worthington, 1968; Freda, 1983; Harris, 1984; Nyman, 1991; Talentino and Landre, 1991; Phillips, 1992; see also Petranka, 1998). Survival to metamorphosis is low, usually ≤ 13% of hatched larvae. In some cases, entire cohorts die due to intense predation and wetland drying (Shoop, 1974; Stenhouse, 1985b, 1987; Stangel, 1988; Ireland, 1989; Figiel and Semlitsch, 1990; Rowe and Dunson, 1995).
Larval food requirements: Larvae are gape-limited and size-selective feeders, ingesting a range of aquatic invertebrates including zooplankton, coleopterans, isopods, ostracods, odonates, and trichopterans. Smaller larvae feed primarily on zooplankton; as they grow in size, consumption shifts towards larger prey such as chironomids, chaoborids, and isopods (Nyman, 1991). Branch and Altig (1981) and Freda (1983) reported that the predominant prey items of larvae were cladocerans and copepods. However, Harris (1995) observed a higher reduction of isopods and amphipods relative to zooplankton under experimental conditions. Freda (1983) also described stomach contents containing eastern newt (Notophthalmus viridescens) larvae with mean SVL 63% of the body size of the larval spotted salamander predators.
Cover: In the presence of predators, refugia such as submerged vegetation are necessary for larval survivorship (Walls, 1995). Spotted salamanders are vulnerable to predation by other ambystomatids, such as marbled salamanders (A. opacum). Interestingly, while spotted salamander larvae require cover, their use of cover does not increase in the presence of competing ambystomatids (Walls, 1995) or fishes (Ireland, 1989; but see Figiel and Semlitsch, 1990). In contrast, Brodman et al. (2002) found that the presence of other Ambystoma caused spotted salamander larvae to occupy refuges more and decrease their activity, and in the presence of A. laterale (blue-spotted salamanders; a potential competitor), to change their activity and use of microhabitat in opposite directions from that of its congener. Stratification (floating in the water column, usually at night) is present in some populations, although its function is not well understood (but see Lannoo and Bachmann, 1984b). Branch and Altig (1981) reported larval stratification in Mississippi ponds. In New Jersey, small larvae tend to remain in leaf litter, while larger larvae stratify in the water column (Nyman, 1991; but see Anderson and Graham, 1967).
Larval polymorphisms Have not been described for spotted salamanders. Given the widespread distribution and numerous population studies on this species, it is unlikely that polymorphisms exist.
Features of metamorphosis: Timing of metamorphosis varies greatly, even within populations, and larval overwintering occurs in some populations that breed in permanent ponds (Phillips and Sexton, 1989; Whitford and Vinegar, 1966). Metamorphosis typically occurs 8–16 wk after hatching, when larvae reach 27.0–32.0 mm SVL (Worthington, 1968; Talentino and Landre, 1991). In many populations that breed in seasonal and semipermanent wetlands, larval survivorship is correlated with wetland duration (Ball, 1937; Ling et al., 1986; Stangel, 1988; see also Petranka, 1998). Abundance and size at metamorphosis are also positively correlated with length of the hydroperiod (Rowe and Dunson, 1995; Phillips et al., 2002). However, in a short hydroperiod year, size at metamorphosis is negatively correlated with time to emergence from the pond, suggesting that faster growing larvae reach an optimal size when compared with other larvae that take longer to metamorphose at smaller sizes (Phillips et al., 2002). Likewise, mean size at metamorphosis in a year when hydroperiod is shorter than usual is significantly smaller than mean size in a longer hydroperiod year (Phillips et al., 2002). At high densities or in the presence of other ambystomatid larvae, spotted salamander larvae have higher mortality, grow more slowly, and metamorphose at smaller sizes (Wilbur, 1972; Wilbur and Collins, 1973). At a Massachusetts pond, larvae that remain in their natal pond longer metamorphose at greater weights than earlier metamorphosing individuals (Windmiller, 1996).
Post-metamorphic migrations: Newly metamorphosed animals dispersing from breeding ponds move beneath rocks and logs near the pond margin where they are sometimes exposed to high temperatures (Pough and Wilson, 1970) and desiccation (Shoop, 1974). Because metamorphosis varies among individuals, the seasonal migration of juveniles away from the ponds is not as synchronized as the adult breeding migrations. Wilson (1976) captured newly metamorphosed animals exiting a New York pond over a period of 5 wk from 27 July–29 August. He recorded sporadic movements ≤ 25 m a night over the course of several nights. In Massachusetts, Windmiller (1996) recorded episodic dispersal of post-metamorphic animals from 1 July to autumn. However, there is one report of a concentration of “hundreds” of juveniles migrating along a dirt road adjacent to a wooded area after a summer storm in Maryland (Hardy, 1952).
Orientation of post-metamorphic individuals in five of six cohorts was highly non-random and biased towards a northward direction in five Massachusetts ponds (Windmiller, 1996). These orientation patterns seemed to reflect the emigration routes of post-breeding parental cohorts and the location of upland forest habitat. In the one post-metamorphic cohort that did not disperse in a northward direction, no forested land occurred to the north of the pond. Most individuals (50%) remained within 40 m of their natal pond as late as 8 December, while 36% were between 40–100 m and 14% dispersed beyond 100 m. Later-metamorphosing individuals were heavier in weight and dispersed on average 18 m farther than those metamorphosing earlier (Windmiller, 1996).
Neoteny: There are no reports of non-transforming individuals in spotted salamanders. Given the widespread distribution and numerous population studies on this species, it is unlikely that neotenic adults exist.
Juvenile Habitat: Habitat characteristics of juveniles are similar to those of adults. Juvenile abundance declines sharply across a gradient running from relatively mature forest to interior habitat (70–90 yr old) to recently clearcut habitat (2–11 yr old) in Maine (deMaynadier and Hunter, 1999). Juveniles are found more frequently closer to the natal pond edge (< 75 m) and seek refuge in burrows and under rocks and fallen logs (Wilson, 1976; Windmiller, 1996). Although usually found only in moist forest habitats, some juveniles were collected from under driftwood on the sandy shore of the Chesapeake Bay in Maryland (Hardy, 1952). Juveniles typically emerge at times beginning in midsummer (Parmalee, 1993). Detailed data for the period of juvenile development are sparse, but presumably they forage on forest floor invertebrates in underground burrows while they mature.
Adult Habitat: Spotted salamanders occur within eastern mixed-deciduous forest habitats and are common in mesic to floodplain habitats. They primarily rely on cutaneous respiration (Whitford and Hutchison, 1966a) and are more commonly found under large, moist cover objects (Parmalee, 1993). As with juveniles, adult abundance declines sharply from relatively mature forest to interior habitat to recently clearcut habitat in Maine (deMaynadier and Hunter, 1999). Along the Atlantic Coastal Plain, spotted salamanders are not common in the drier, sandy, upland environments and are mostly found in hardwood, bottomland habitats with lower soil temperatures and higher soil moisture. These habitats frequently harbor fish populations that significantly decrease the reproductive success of spotted salamanders but do not exclude them from breeding there (Semlitsch, 1988). Potential habitat has been described by Baldwin and Vasconcelos (2003).
Home Range Size: Burrow systems of individual adults in Kentucky and Michigan encompass 12–14 m2 and an average of 9.8 m2 of forest floor, respectively (Douglas and Monroe, 1981; Kleeberger and Werner, 1983). In Michigan, average home range movements of 14 ± 3 m were recorded over a 6–7 wk period. During this time, adults were mostly found in burrows (72% of the time), but also in decaying logs (21%) and leaf litter (7%; Kleeberger and Werner, 1983).
Territories: Although territoriality has not been documented in larval salamanders, larvae exhibit visual and movement displays that function as aggressive behavior in crowded environments (Walls and Semlitsch, 1991). In the larval stage, refuge use and stratification may minimize such aggressive interactions through habitat partitioning (Branch and Altig, 1981; Nyman, 1991). Adults are almost always the sole residents under cover objects and are rarely found in close proximity in their forest habitats (Parmelee, 1993). In laboratory trials, adults will alter their normal movement patterns to avoid conspecifics (Ducey and Ritsema, 1988). In other laboratory trials, adults actively defended experimental burrows and feeding areas against conspecific and heterospecific intruders (Ducey and Ritsema, 1988; Walls, 1990; Ducey and Heuer, 1991). Conspecific interactions involve biting, head butting, and forebody raising (Ducey and Ritsema, 1988). In one case of interspecific interactions, spotted salamanders consumed 9% of an eastern red-backed salamander (Plethodon cinereus), but this aggression may have been due to predation rather than territorial defense (Ducey et al., 1994).
Aestivation/Avoiding Dessication: In some areas of the range, adults are likely to experience surface temperatures above their critical thermal maximum (34.9–35.8 ˚C; Gatz, 1971, 1973b) and will usually seek refuge from heat and desiccation in small mammal burrows. Adults have been found in burrows as deep as 1.3 m (Semlitsch, 1983c).
Seasonal Migrations: Adults move to and from breeding ponds (see "Breeding migrations" above). Post-metamorphic animals migrate away from wetlands from mid to late summer (see Faccio, 2003).
Torpor (Hibernation): Adults and post-metamorphic animals hibernate in subterranean burrows and likely keep from freezing by moving below the frost line.
Interspecific Associations/Exclusions: The structure of larval communities has been the focus of numerous studies on the effects of phenology, density, and interspecific competition among sympatric ambystomatids. Spotted salamanders often breed syntopically with congeners, including Jefferson salamanders (A. jeffersonianum), marbled salamanders, mole salamanders (A. talpoideum), eastern tiger salamanders (A. t. tigrinum), blue-spotted salamanders, and small-mouthed salamanders (A. texanum). Many studies have investigated the mechanisms of coexistence among these species, and in general, spotted salamander larvae are inferior direct competitors because they generally are smaller than their interspecific competitors. However, female Cope's gray treefrogs (Hyla chrysoscelis) avoid depositing eggs in pools containing larval spotted salamanders, suggesting that larvae prey on hatchling tadpoles (Resetarits and Wilbur, 1989).
Brodman (1995, 1996) found that density of each species was an important component determining the outcome of interactions with conspecific and heterospecific competitors. Species show negative responses, such as reduced growth rate and smaller body size at metamorphosis to increased density (Semlitsch and Walls, 1993). Walls and Jaeger (1987) and Walls (1996) demonstrated that spotted salamander larvae were superior exploitative competitors when in competition with mole salamanders. However, Brodman (1999b) argued that coexistence between spotted salamanders and Jefferson salamanders could not be explained by the interference-exploitative competition trade-off, because spotted salamanders did not reduce the growth of congeners at high densities. Other factors influence interspecific competition, such as spatial or dietary partitioning through refugium selection (Walls, 1995; Brodman, 1996), temperature preference (Anderson and Graham, 1967; Stauffer et al., 1983), or dietary differences (Nyman, 1991). Larval spotted salamander behavior is highly variable and may explain their success in a variety of competitive larval environments throughout their range (Brodman, 1996). For example, spotted salamanders reduce the amount of time they spend in the water column (stratification) in the presence of interspecific predators (Walls and Williams, 2001). Predator-mediated reduction in activity patterns potentially limits foraging opportunities and hence potentially decreases survival in this species. Thus, in some cases the composition of larval amphibian assemblages could exclude spotted salamanders from communities of pond-breeding amphibians (Walls and Williams, 2001), yet they continue to persist in many highly competitive larval environments.
Juvenile and adult spotted salamanders live under cover objects and in burrows created by shrews, moles, and other small mammals (Gordon, 1968; Douglas and Moore, 1981; Madison, 1997). However, the associations among co-inhabitants of burrow systems are not well known. Laboratory experiments show that juvenile spotted salamanders exhibit preference for unoccupied burrows and will leave burrows already inhabited by conspecific or heterospecific (marbled salamander) juveniles (Smyers et al., 2002). Adults also share home ranges with other salamanders, including eastern red-backed salamanders (Ducey et al., 1994) and mole salamanders (Walls, 1990). Spotted salamanders share breeding habitats with numerous species, including fishes. Naive adults do not adjust their breeding behavior in the presence of fish, nor do females reduce the number of eggs they deposit (Sexton et al., 1994). However, larvae do seem to adopt behaviors that protect them from fish predation (Figiel and Semlitsch, 1990).
Age/Size at Reproductive Maturity: Adults generally range in size from 15–25 cm TL (Petranka, 1998). In southern populations, individuals may take < 3 yr to reach sexual maturity (Bishop, 1941b, 1943; Minton, 1972, 2001). In northern populations, males reach maturity earlier than females. In a Québec population examined using skeletochronology, the majority of animals were 2–18 yr old with the mode at 6–8 yr. Most females matured by 7 yr at ≥ 78 mm SVL. Males reached maturity between the ages of 2 and 6 and at ≥ 63 mm SVL (Flageole and Leclair, 1992). In Michigan populations, males reach sexual maturity in 2–3 yr and females in 3–5 yr (Wilbur, 1977c; see also Petranka, 1998). In Massachusetts, some males attain sexual maturity in 2 yr, while females require an additional year (Windmiller, 1996). At this locality, the smallest sexually mature individuals captured were 56.5 and 60.2 mm SVL for males and females, respectively. Growth in spotted salamanders has been analyzed by Blackwell et al. (2003) and Homan et al. (2003).
Longevity: Individuals live ≤ 22–25 yr in captivity (P.H. Pope, 1928a, 1937; Snider and Bowler, 1992), and skeletochronology indicated the oldest individual was 32 yr in a Québec population (Flageole and Leclair, 1992). Several mark-recapture studies at breeding ponds suggest that adult survivorship is high from one breeding season to the next, ranging from 70–100% (Husting, 1965; Whitford and Vinegar, 1966; Douglas and Monroe, 1981; see also Petranka, 1998). Females appear to suffer higher mortality than males (Harris, 1980).
Feeding Behavior: During the breeding season adults apparently do not feed (Smallwood, 1928). In terrestrial sites, adults and juvenile spotted salamanders feed by either protruding their heads from burrows using a sit-and-wait strategy or by foraging more actively in wet weather on the forest floor. They are generalists on forest floor invertebrates, consuming mollusks, earthworms, centipedes, millipedes, spiders, and a wide variety of insects (Bishop, 1941b; Pope, 1944). Given the amount of time adults spend in burrows and the depths at which they are found (1.3 m; Gordon, 1968; Semlitsch, 1983c), they may also feed underground.
Predators: Larval wood frogs (Rana sylvatica), centrarchid and cyprinid fishes, and various larval invertebrate species prey on spotted salamander eggs (Ward and Sexton, 1981; Semlitsch, 1988; Petranka, 1998; Baldwin and Calhoun, 2002). In a Virginia population, high reproductive failure was attributed to predation by centrarchid fishes (Ireland, 1989). Eastern newt adults will eat spotted salamander eggs (Hamilton, 1932; Wood and Goodwin, 1954), as will caddisfly and midge (Parachironomus sp.) larvae (Murphy, 1961; Leclair and Bourassa, 1981; Stout et al., 1992).
Larvae are preyed upon by caddisflies (Ptilostomis postica; Murphy, 1961; Rowe and Dunson, 1993; Rowe et al., 1994; and Banksiola dossuaria; Stout et al., 1992) and other predatory aquatic insects such as midge (Parachironomus sp.) larvae (Petranka, 1998). Wood frog tadpoles will prey on larvae (Petranka, 1998; Petranka et al., 1998; Baldwin and Calhoun, 2002), as will least sandpipers (Calidris minutilla; Stangel, 1983). Larval predators also include other Ambystoma species; in regions of sympatry, marbled salamander larvae are already present and will prey on spotted salamander larvae when they hatch (Stewart, 1956; Husting, 1965; Stenhouse, 1985b). Thus, it is not surprising that spotted salamander larval densities are inversely correlated with marbled salamander larval densities (but see Worthington, 1968; Cortwright, 1988; Petranka, 1998). Jefferson salamander and silvery salamander (A. platineum) larvae also prey on spotted salamander larvae (Noble, 1931; Nyman, 1991; Brodman, 1996). Newly metamorphosed animals and breeding adults are preyed on by raccoons (Huheey and Stupka, 1967) and probably other mammals such as opossums, weasels, and minks (Beachy, 1991a).
Anti-Predator Mechanisms: Egg masses have a thick jelly coat that may afford some protection from centrarchid fishes and other aquatic invertebrate predators (Ward and Sexton, 1981; Semlitsch, 1988). There are conflicting reports on whether or not leeches prey on eggs (Cory and Manion, 1953; Cargo, 1960). Cunnington and Brooks (2000) tested whether adults breeding in harsher environments containing predatory fishes produced larger eggs—which might offer some protection to larvae against predation—but found no significant differences. Larvae possess no direct anti-predator mechanisms (e.g., Anderson and Petranka, 2003) but are less prone to predation when refuges are available. One experiment of microhabitat partitioning demonstrated that spotted salamander larvae tend to occupy refuges at a higher rate and with lower activity levels in the presence of predators (Brodman et al., 2002).
Spotted salamanders behave aggressively towards potential predators, head butting and biting attackers (Dodd, 1977b; Brodie et al., 1979). Adults can assume defensive postures, vocalize, and produce noxious skin secretions from granular glands located primarily on the dorsolateral surface of the tail and the paratoid gland on the head (Barach, 1951; Howard, 1971; Wyman and Thrall, 1972; Brodie, 1978; Brodie et al., 1979). These secretions are effective deterrents to predation by short-tailed shrews (Blarina brevicauda; Brodie et al., 1979) and potentially against some reptilian predators (Barach, 1951; Howard, 1971), presumably because they are distasteful. Brodie (1971a) reported that the secretions “cause a drying and burning sensation to the tongue." Nonetheless, adult spotted salamanders have been found in the stomachs of common garter snakes (Thamnophis sirtalis, Klemens, 1993), eastern hog-nosed snakes (Heterodon platirhinos; Babbitt, 1932), and fishes (Salmo trutta; Bishop, 1941b).
Diseases: In egg clutches collected from several Michigan ponds and raised in the laboratory, protozoans (Tetrahymena sp.) were found to infect and kill the developing embryos (Ling and Werner, 1988; see also Petranka, 1998). Evidence of infection was characterized by swelling of the epidermis, and histological preparations revealed Tetrahymena sp. concentrations in nervous tissue. Corliss (1954) originally found Tetrahymena sp. in egg masses and developing embryos, but no effects were noted. Fungal infections have been observed in egg masses maintained in the laboratory, as well as in nature, but have only been mentioned anecdotally in the literature (e.g., Ward and Sexton, 1981).
Kingsley (1880) first reported the occurrence of natural limb anomalies in spotted salamanders. Polydactyly (particularly of the forelimbs), syndactyly of the proximal first digits, and missing digits were common abnormalities observed in juveniles and adults in a New York population (Wilson, 1976). Cases of melanism, albinism, and partial albinism are documented in adults, but usually occur in populations at low frequencies (Brandt, 1952; Hensley, 1959; Smith and Michener, 1962; Husting, 1965; Easterla, 1968; Mount, 1975; Dyrkacz, 1981). In a Mississippi population, Worthington (1974) found a high incidence of abnormal coloration, deviant counts of vertebral trunk elements, and limb anomalies; he suggested high temperatures as a cause, although genetic and other environmental factors could not be ruled out.
Parasites: Rankin (1937) reported the following parasites from spotted salamanders in North Carolina: Protozoa—Cytamoeba bactifera, Euglenamorpha hegneri, Hexamitus intestinalis, Prowazekella longifilis, Tritrichomonas augusta; Trematoda—Brachycoelium hospitale, Diplostomulum ambystomae; Nematoda—spirurid cysts; and Acarina—Hannemania dunni.
Bolek and Coggins (1998) reported helminth species infecting the small and large intestine of spotted salamanders from northwest Wisconsin. Of 20 individuals sampled, 9 were infected by the nematode Batracholandros magnavulvaris; the trematode Brachycoelium salamandrae was found in only three individuals but at a higher mean intensity. Kuzmin et al. (2001) also described a nematode in the lungs and body cavity of spotted salamanders from northwestern Wisconsin; this nematode, Rhabdias ambystomae, is the first species of the genus described from salamanders in North America. Leeches have also been commonly observed on adults within and exiting breeding ponds. The impact, if any, of helminth or leech parasitism on spotted salamander population dynamics has not been documented.
Conservation: Spotted salamander populations are seemingly widespread throughout the eastern United States, frequently occurring in nature preserves and state and federal parks, as well as in relatively undisturbed forests and private lands. Although present in fragmented suburban areas (Klemens, 1993; Gibbs, 1998a; Wright and Zamudio, 2002) and sometimes in highly disturbed urban areas (Klemens, 1993; DiMauro and Hunter, 2002), spotted salamander populations decline with increasing urbanization (Windmiller, 1996) and mere presence indicates little about long-term population viability in these habitats. Because populations are vulnerable to human developments that alter or eliminate habitats and corridors, they are at risk of local extinction or even local population genetic consequences. Population comparisons using fluctuating asymmetry in dorsal spot patterns as a correlate of habitat disturbance showed that a population of breeding adults was significantly more asymmetrical in a disturbed landscape (in this case, a golf course) than in a nature reserve (Wright and Zamudio, 2002). These findings suggest that populations in the disturbed habitats may experience reductions in fitness, potentially due to demographic/genetic bottlenecks or developmental perturbations (due to chemical applications) as measured by fluctuating asymmetry, despite their continued persistence in the disturbed habitat.
Connectivity among patches of breeding habitat will decrease with habitat alteration. Thus, the importance of individual populations increases because viable populations can rescue nearby sinks in a metapopulation framework. Semlitsch and Bodie (1998) examined frequency distribution of wetland sizes and concluded that even small, isolated wetlands serve as important sources of juvenile recruits. Their loss could result in a direct reduction of connectivity among remaining populations. Spotted salamanders are likely to benefit from efforts to maintain connectivity among patches of upland forest habitats and aquatic breeding sites with sufficient buffer zones (Semlitsch, 1998). Spotted salamanders are more likely to occupy ponds in more forested areas and those that are adjacent to forest (Guerry and Hunter, 2002). Thus, the composition and configuration of the landscape surrounding breeding ponds is associated with the likelihood of spotted salamander presence and population persistence.
Environmental contamination in breeding ponds may negatively affect juvenile recruitment in spotted salamanders. Habitat acidification is a concern for this species, as hatching success and larval development can be reduced in pH ranges of 4.5–5.5 (Pough, 1976; Pough and Wilson, 1977; Ling et al., 1986; Sadinski and Dunson, 1992; Brodman, 1993), levels that are not uncommon in the eastern United States (Clark, 1986b; Cook, 1983; Ling et al., 1986; Sadinski and Dunson, 1992; Petranka, 1998). The number of eggs present in a pond is positively correlated with alkalinity, as is hatching success (Clark, 1986b; Petranka, 1998). Ireland (1991) demonstrated that significantly reduced growth rates were not the result of pH alone but may be a function of anion concentration or the combined effect of anion and hydrogen ions. Cook (1983) found no correlation between low pH and embryonic mortality in 13 Massachusetts ponds; in contrast, Portnoy (1990) found reduced survivorship correlated with acidification in the Cape Cod region.
Low pH levels can also reduce growth and survival rates, as well as alter competitive interactions between larval spotted salamanders and Jefferson salamanders (Brodman, 1993). Acidification is not the only threat to adult reproductive fitness. Over an 8-yr period in eastern Virginia, reproductive activity of spotted salamanders declined severely in breeding ponds with low pH levels and high aluminum, copper, silicon, and zinc concentrations (Blem and Blem, 1989, 1991). Elevated hydrogen ion and aluminum concentrations in breeding ponds may also negatively affect hatching success (Clark and Hall, 1985). In Pennsylvania, the number of eggs present is positively correlated with pH and pond size, but negatively correlated with cations and silica levels (Rowe and Dunson, 1993).
During the breeding season, road mortality is a considerable threat to migrating amphibians. Although the direct effects of road mortality on population numbers have not been thoroughly evaluated, several studies show that amphibian densities decrease in response to roadways (Fahrig et al., 1995; Vos and Chardon, 1998; Trombulak and Frissell, 2000). The direct effects of roads on mortality may not be the only consideration. In New Hampshire, embryonic survivorship is reduced in breeding ponds contaminated with road salt runoff (Turtle, 2000). Roads also affect animal communities by creating habitat edges that promote invasion of exotic plants (Gaddy and Kohlstaat, 1987) and exposing salamanders to predation. Moreover, heavily trafficked roads may entirely block migrating salamanders from reaching a breeding pond (Trombulak and Frissell, 2000). In Massachusetts, Jackson and Tyning (1989) reported that drift fences and two tunnels installed under a road aided migrating spotted salamanders and substantially reduced mortality. Migratory routes under roads are not difficult to implement, and amphibian populations would benefit from local efforts to reduce road mortality during their short breeding seasons.
References
- Altig, Ronald & McDiarmid, Roy W. 2015. Handbook of Larval Amphibians of the United States and Canada. Cornell University Press, Ithaca, NY. 341 pages.
- AmphibiaWeb. 2020. University of California, Berkeley, CA, USA.
- Conant, R., 1975, A field guide to reptiles and amphibians of Eastern and Central North America, 429 pgs., Houghton Mifflin Co., Boston, MA
- Bishop, S.C., 1943, Handbook of Salamanders, 555 pgs., Comstock Publ. Co., New York, NY
- Petranka, J.W., 1998, Salamanders of the United States and Canada, 587 pp. pgs., Smithsonian Institution Press, Washington, DC
Photos:
*Click on a thumbnail for a larger version.

 Metamorph Arlington _small.jpg)





 larvae.jpg)


Verified County/City Occurrence
Accomack
Albemarle
Alleghany
Amelia
Amherst
Appomattox
Arlington
Augusta
Bath
Bedford
Botetourt
Brunswick
Buckingham
Campbell
Caroline
Carroll
Charles City
Charlotte
Chesterfield
Craig
Culpeper
Cumberland
Dinwiddie
Essex
Fairfax
Fauquier
Floyd
Fluvanna
Franklin
Frederick
Giles
Goochland
Greensville
Halifax
Hanover
Henrico
Henry
Highland
Isle of Wight
James City
King George
Lancaster
Lee
Loudoun
Louisa
Lunenburg
Madison
Mecklenburg
Middlesex
Montgomery
Nelson
New Kent
Northampton
Northumberland
Nottoway
Orange
Page
Patrick
Pittsylvania
Powhatan
Prince Edward
Prince George
Prince William
Pulaski
Rappahannock
Richmond
Roanoke
Rockbridge
Rockingham
Russell
Shenandoah
Smyth
Southampton
Spotsylvania
Stafford
Surry
Sussex
Warren
Washington
Westmoreland
Wise
Wythe
York
CITIES
Charlottesville
Danville
Harrisonburg
Lynchburg
Martinsville
Richmond
Waynesboro
Williamsburg
Verified in 83 counties and 8 cities.
U.S. Range

Box Turtle Reporting
