Northern Scarletsnake
Cemophora coccinea copei
** Harmless **
Common Name: |
Northern Scarletsnake |
Scientific Name: |
Cemophora coccinea copei |
Etymology: |
|
Genus: |
Cemophora is derived from the Greek words kemos meaning "muzzle" and phoreus meaning "bearer". |
Species: |
coccinea is derived from the Latin word coccineus which means "scarlet" or "crimson". |
Subspecies: |
copei is a patronym honoring Edward Drinker Cope. |
Vernacular Names: |
False coral snake, milk snake |
Average Length: |
14 - 20 in. (36 - 51 cm) |
Virginia Record Length: |
25.6 in. (65.1 cm) |
Record length: |
32.6 in. (82.8 cm) |
Systematics: Originally described as Coluber coccineus by Johann Friedrich Blumenbach in 1788. He designated no type specimen, but indicated "Florida" as the type locality. A more restricted type locality has not been specified. Cope (1860) was the first to use the generic name Cemophora for this species. Two subspecies are recognized: C. coccinea copei and C. coccinea coccinea. Their distributions were illustrated in Williams (1985) and Conant and Collins (1991). Only C. coccinea copei occurs in Virginia. This subspecies was first described as Cemophora copei by Giorgio Jan in 1863 based on a specimen from "Etats-Unis, Tennessee." Garman (1883) was the first to consider it a subspecies of C. coccinea. All Virginia authors have used the current nomenclature.
Description: A colorful, moderate-sized snake reaching a maximum total length of 828 mm (32.6 inches) (Conant and Collins, 1991). In Virginia, maximum known snout-vent length (SVL) is 575 mm (22.6 in.) and maximum total length is 651 mm (25.6 in.). In this study, tail length/total length averaged 13.5 ± 1.3% (11.3-16.3, n = 26).
Scutellation: Ventrals 158-174 (ave. = 161.9 ± 3.9, n = 28); subcaudals 33-48 (ave. = 39.0 ± 3.5, n = 28); ventrals + subcaudals 192-214 (ave. = 200.9 ± 5.0, n = 28); dorsal scales smooth, scale rows usually 19 (80%, n = 24) at midbody, but may be 20-21 (20%); anal plate undivided; infralabials usually 7/7 (61.9%, n = 21) or other combinations of 6-8 (38.1%); supralabials usually 6/6 (69.6%, n = 23) or other combinations of 5-6 (30.4%); loreal present; preoculars 1/1; postoculars 2/2; temporals 1+2/1+2 (100%, n = 23); rostral scale enlarged.
Coloration and Pattern: Dorsal pattern consists of a series of 13-20 red blotches (ave. = 16.6 ± 2.1, n = 27) on body and 3-6 on tail, each of which is strongly outlined in black anteriorly and posteriorly; black pigment on lateral edges irregular to faint; first black body saddle touches black head band or is separated from it by 1 scale width; red portion of anteriormost saddle 5-9 scales in length and black borders 1-2 scales wide; mid-body red saddle 3-8 scales long and black borders 1-2.5 scales wide; black pigment of saddles encroaches onto lateral edges of ventral scales in some individuals; spaces between saddles pale yellow or white and may be peppered with black to brown pigment in large individuals; these spaces 0.5-2 scales wide at midbody; venter and lower jaw cream in color and immaculate; anterior portion of head red; a black band lies across head behind eyes. A muzzlelike snout formed from the enlarged rostral scale projects beyond the lower jaw. The head is slightly elongated and is only as wide as the neck. The last 2 rear teeth on the maxillary bones in the upper jaw are enlarged and saberlike.
Sexual Dimorphism: Average adult SVL was greater in males (450.1 ± 99.6 mm, 295-575, n = 9) than in females (361.9 ± 76.7 mm, 260-483, n = 9). Sexual dimorphism index was -0.24. Body mass is unknown. Males and females were similar in average number of ventral scales (males 161.3 ± 3.3, 158-168, n = 12; females 162.4 ± 4.3,158-174, n = 16), but males had a higher average number of subcaudal scales (41.3 ± 3.0, 36-48, n = 12) than females (37.2 ± 2.7, 33-42, n = 16). No sexual dimorphism was exhibited by average tail length/ total length (males 13.5 ± 1.3%, 11.5-15.2, n = 11; females 13.4 ± 1.3%, 11.3-16.3, n = 15) or in average number of body blotches (males 16.7 ± 2.6, 14-19, n = 12; females 16.6 ± 1.5, 13-20, n = 15). The average number of ventrals + subcaudals was only slightly higher in males (202.6 ± 5.3, 194-214, n = 12) than females (199.6 ± 4.5, 192-209, n = 16). There were no apparent differences in color and pattern.
Juveniles: Juveniles at hatching are patterned as adults but the saddles are pinkish rather than red (Woolcott, 1959). The colors become darker and duller in old individuals. Hatchlings from a single Virginia female averaged 156 mm (150-164) total length (Woolcott, 1959).
Confusing Species: In Virginia, this species may be confused with Lampropeltis triangulum triangulum or Lampropeltis elapsoides.
Geographic Variations: Variation in scutellation and pattern is not pronounced in Virginia. Specimens from the Coastal Plain had 192-214 ventrals + subcaudals (ave. = 201.7 ± 5.4, n = 19), whereas those from the Piedmont had 195-205 (ave. = 199.5 ± 3.4, n = 8). Average number of body blotches was slightly fewer in the Coastal Plain (13-20, ave. = 16.2 ± 2.0, n = 19) than in the Piedmont (15-20, ave. = 17.6 ± 2.0, n = 8). Williams and Wilson (1967) described geographic variation throughout the range and demonstrated a clinal increase in ventral scales and number of body blotches from New Jersey to the lower Mississippi states.
Biology: Northern Scarletsnakes are found in areas where the soil is loose, well drained, and (usually) sandy, and where the vegetation is dominated by pine trees. This snake is a burrower, seldom found in day-light except in or under logs and other surface objects; most have been encountered in late spring or summer as they crossed paved roads at night. All Virginia C. coccinea were captured between 19 April and 7 November. Specimens caught by Gibbons and Semlitsch (1991) in South Carolina were found April-September.
Northern Scarletsnakes are reported to eat skinks (Plestiodon, Scincella), small snakes, frogs, small mice, and soft-bodied insects, although reptile eggs, particularly eggs of small snakes, are preferred (Williams and Wilson, 1967; Palmer and Tregembo, 1970). Burger et al. (1992) observed an individual C. coccinea eating a Northern Pinesnake (Pituophis melanoleucus) egg in New Jersey; its head was inside. Eggs are slit with the enlarged maxillary teeth and the contents squeezed into the mouth with a body coil and the snake's chewing action (Palmer and Tregembo, 1970). Other prey are constricted. Of 10 Virginia snakes examined, four unidentifiable eggs were found in one 536-mm-SVL adult male and a worm was found in a 118-mm-SVL juvenile. Brown (1979) noted that a large Southern Toad (Bufo terrestris) partially swallowed a C. coccinea in South Carolina. Females sometimes eat their own eggs or the eggs of conspecifics (Wright and Wright, 1957; Palmer and Tregembo, 1970).
Woolcott (1959) reported the only known information on reproduction of C. coccinea in Virginia. On 12 September 1956 he found a clutch of five eggs under moist humus in pine woods in Henrico County averaging 37 mm X 17 mm and weighing an average of 5.9 g. They hatched on 25 September. Clutches of 2-9 eggs (ave. = 5.8) were reported by Ernst and Barbour (1989a) for this species. The smallest mature male I examined was 295 mm SVL and the smallest mature female was 260 mm SVL. Gibbons and Semlitsch (1991) reported finding a juvenile in May emerging from an egg that was in sandy soil 1.3 m below the surface. Do C. coccinea hatchlings overwinter in the nest?
Northern Scarletsnakes are one of the few in Virginia that actively burrow in soil. The head is thrust forward, upward, and side to side while forcing the snout deeper into the soil (Williams and Wilson, 1967). Nelson and Gibbons (1972) reported that C. coccinea moved at minimal rates of 15 m per day during its active season. Clifford (1976) listed only one C. coccinea among the 278 snakes found in a 4-year period in Amelia County. The population ecology of this species is unknown.
Remarks: Other common names in Virginia are false coral snake and candy cane snake (Linzey and Clifford, 1981).
Due to the superficial resemblance of Cemophora coccinea to coral snakes (Micrurus fulvius) in south-eastern North America, they are sometimes called coral snake mimics. Mimicry conveys some protection to the mimic from visual predators because these predators learn that the model, the coral snake in this case, is distasteful or harmful. That the color and pattern in scarlet snakes evolved because it conveyed protection as mimics of coral snakes has yet to be proven, however.
Due to the dearth of information available, all observations on the biology of this secretive snake should be reported.
Conservation and Management: In the first attempt to recognize species of special concern, Russ (1973) listed this snake in the status undetermined category. Cemophora coccinea appears to be secure in most areas of Virginia, although habitat loss has become a threat in areas of rapid urbanization. Information on local population sizes and dynamics, as well as abundance in various habitat types, is needed for the development of realistic management strategies. The continued existence of northern Virginia populations needs to be verified as extensive urban development has eliminated much of the habitat. The introduced red fire ant (Solenopsis invicta) may become a threat to some populations.
References for Life History
Photos:
*Click on a thumbnail for a larger version.




Verified County/City Occurrence
Alleghany
Amelia
Brunswick
Caroline
Charles City
Charlotte
Chesterfield
Cumberland
Dinwiddie
Gloucester
Hanover
Henrico
Henry
Isle of Wight
James City
King and Queen
New Kent
Nottoway
Powhatan
Prince Edward
Prince George
Southampton
Surry
Sussex
CITIES
Chesapeake
Fairfax
Newport News
Petersburg
Virginia Beach
Verified in 24 counties and 5 cities.
U.S. Range
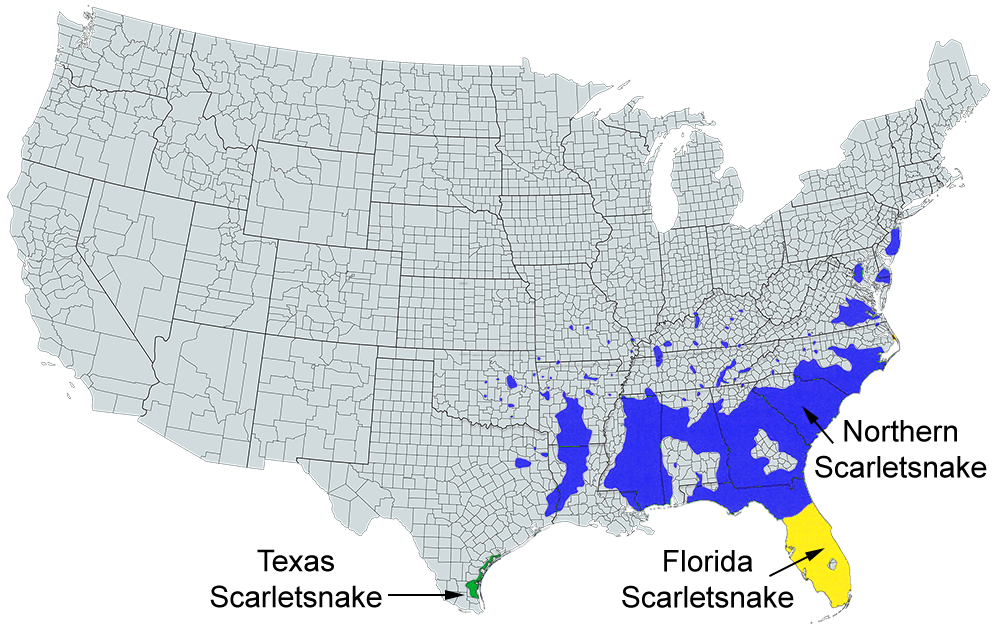
US range map based on work done by The Center for North American Herpetology (cnah.org) and Travis W. Taggart.

Box Turtle Reporting
